Определение пола и возраста воробьиных птиц фауны СССР, методика

На основании большого опыта авторов по определению пола и возраста птиц при кольцевании, полного просмотра коллекций Зоологического института АН СССР и некоторых других коллекций, обнаружено много новых и проверены ранее известные способы определения пола и возраста у живых птиц. Справочник состоит из трех разделов: подробного описания методов поиска признаков пола и возраста у малоизученных видов, описания методик прижизненного анализа отлавливаемых для кольцевания птиц, и описания в систематическом порядке признаков подвидовой принадлежности (в отношении подвидов, опознаваемых по единичной особи), пола и возраста всех воробьиных птиц фауны СССР.
ПРЕДИСЛОВИЕ
В последние годы в СССР началось быстрое развитие кольцевания птиц, которое будет все увеличиваться в ближайшие годы - охватывая новые виды и неизученные территории. Необходимо, чтобы данные о каждой окольцованной птице были точными и полными. Только в таком случае накопление находок окольцованных птиц позволит в будущем полноценно их анализировать. В первую очередь нужно стремиться к тому, чтобы у кольцуемых птиц были определены пол и возраст. Кольцевание само по себе — деятельность с низкой результативностью и большим запаздыванием в получении результатов, особенно если вид дает низкий процент возвратов или работа проводится на малонаселенной территории. Но эффективность работы резко меняется, если кольцевание сопровождается прижизненной обработкой кольцуемых птиц, которая сразу дает массовый ценный материал о свободноживущих птицах. Поэтому кольцевание птиц целесообразно сопровождать прижизненным анализом их, проводимым по стандартной системе. Все это требует справочников и руководств по кольцеванию, определению пола и возраста и прижизненному анализу птиц, адресованных кольцующему орнитологу и приспособленных к специфике работы с живыми птицами.
Разработка таких пособий в некоторых странах началась еще в тридцатые годы (Drost, 1930—1940); к нашему времени вышло несколько руководств для фаун стран Европы (Arnheim J. et R., 1968; Drost, 1951; Cornwallis, Smith, 1960a), отдельных групп птиц (Williamson, 1960, 1962, 1964) и, наконец, для всей Европы (Svensson, 1970). Для фауны СССР подобное руководство публикуется впервые.
Настоящий справочник состоит из нескольких разделов, преследующих разные, но взаимосвязанные цели.
1. Приведены методы нахождения признаков пола и возраста у живых воробьиных птиц. Для многих птиц нашей фауны эту работу еще следует осуществить, и сделать это могут только активно кольцующие орнитологи, получающие в свои руки большое количество живых птиц.
2. Изложены проверенные и оправдавшие себя методы прижизненного анализа состояния птиц и способы последующей аналитической обработки собранного первичного материала
3. Дано повидовое руководство для определения пола и возраста воробьиных птиц нашей фауны путем прижизненного осмотра. Признаки приводятся раздельно для разных сезонов, в том числе и для миграции.
4. Справочник одновременно указывает степень изученности этих признаков для разных видов и дает дополнительные сведения справочного характера по размерам птиц, их весу и характеру линьки.
ОБЩАЯ ЧАСТЬ
ПРАВИЛА КОЛЬЦЕВАНИЯ
Метод предполагает, что из двух человек, участвующих в добывании одиночного научного факта, первый, кольцующий, должен быть компетентным орнитологом, а второй — нашедший птицу с кольцом — как правило, некомпетентен. Поэтому на кольцующем лежит вся ответственность за качество результатов кольцевания. Кольцевание — коллективная деятельность многих поколений орнитологов, а его результаты — всемирная коллекция, которая обрабатывалась и будет обрабатываться неоднократно и в разных направлениях. Протоколы кольцевания должны быть заполнены в полном соответствии с требованиями и содержать всю необходимую информацию. Результаты кольцевания считаются научными аргументами высшей ценности, поэтому любая ошибка приносит очень большой вред.
Семьдесят пять лет кольцевания птиц научили, что каждый принимающий в нем участие, независимо от, своих личных целей, должен строго выполнять некоторые перечисленные ниже правила, нарушение которых наносит ущерб науке.
1. Надевание кольца — не первое действие при кольцевании птицы. До этого она должна быть определена и запись должна быть сделана. Если начать с надевания кольца, а птица вырвется и улетит, кольцо будет носить неизвестно кто.
2. Если возникают сомнения в определении вида взрослой птицы или птенцов, их не следует кольцевать.
3. Указывать следует только абсолютно достоверную информацию. Если подвид птицы неясен,— не нужно его указывать. Нельзя писать подвид по соображениям географического положения места кольцевания относительно ареала. Это сделает по тем же соображениям орнитолог, обрабатывающий возвраты. Если нет уверенности в определении пола и возраста птицы, они не указываются. Не следует писать: самка ? sad?- этот вопрос останется навсегда неразрешенным. Лучше в примечании указать признаки, вызвавшие сомнения. В будущем эти признаки могут быть проверены и пол определен по записи.
4. Необходимо следить за соответствием номеров колец на птицах и в протоколе. Кольца часто поступают с нарушением порядка, пропусками и дублями. К тому же они легко теряются, поэтому номер каждого кольца непосредственно перед тем, как надеть на птицу, нужно сверить с номером в записи. Поздно обнаруженная потеря одного кольца приводит к тому, что десятки птиц получили неверную, неисправимую запись.
5. Нужно заполнять все графы протокола, в том числе и те, ответ на которые отрицательный,— это обеспечивает однозначность понимания.
6. Записи следует вести набело. В любых условиях записи нужно делать сразу в стандартный журнал кольцевания. Всякое переписывание, во-первых, приводит к ошибкам, а, во-вторых, часто откладывается на «потом», что в некоторых случаях оказывается «навсегда».
7. Надписи делаются в соответствии с установленной формой: название вида по латыни и целиком, пол — соответствующими значками; возраст — латинскими сокращениями; дату с днем, месяцем и годом; место — согласно официальным административным названиям, с максимально объективной привязкой (азимут и расстояние от легко находимой на карте точки).
ОСМОТР И ИЗМЕРЕНИЕ ЖИВЫХ ПТИЦ, УПОТРЕБЛЯЕМЫЕ ТЕРМИНЫ
С пойманной птицей обращаться нужно бережно и задерживать ее как можно меньше времени. Для переноса и сохранения птиц удобно применять специальные садки, например, такой вариант, как на Биологической станции в Рыбачьем. Садок представляет собой легкий деревянный ящик, высотой 15 X 30 X 60 см, перегороженный на две части. Дно из фанеры, верх из мелкоячеистой (6—8 мм) дели. Садок переносится за ручку из двух диагонально пересекающихся шнуров, прикрепленных в каждом углу. В узких боковых стенках садка прорезаны два отверстия 10 X 10 см, к краям которых пришиты «рукава» из плотной ткани длиной 30 см для просовывания руки в садок. Через них птиц помещают в садок и вынимают. Садок закрывают, проталкивая скомканный рукав внутрь или завязывая его. Нужно иметь несколько таких садков, снабженных номерами и отметкой левого и правого отсеков, чтобы их не путать.
От поимки до выпуска птицы летом должно проходить не более 30 мин. Во время массового отлова на пролете задержка не должна превышать часа. Мелких и крупных, агрессивных и неагрессивных птиц следует помещать в разные садки. В первую очередь обрабатывают более слабых и мелких птиц. Пойманные выводки нужно выпускать целиком; выкармливаемых родителями молодых выпускать на месте поимки. Следует сохранять стаи (т. е. выпускать одновременно) у видов с постоянным составом стай, например Aegithalos caudatus. Если птиц много, целесообразно выпускать их не по одной, а небольшими партиями. Для этого обработанных птиц спускают по наклонной скользкой трубе в низкий плоский ящик. Раз в несколько минут верхнюю крышку ящика откидывают, и птицы вылетают. Для плотностайных птиц это необходимо, чтобы не разбивать сложившуюся стаю. При этом группа птиц немедленно вся улетает хотя бы на некоторое расстояние от места кольцевания, в то время как птицы, выпущенные поодиночке, часто оседают на некоторое время тут же рядом, снова попадаются в ловушки и привлекают хищников, которые легко ловят ошарашенных процедурой кольцевания птиц.
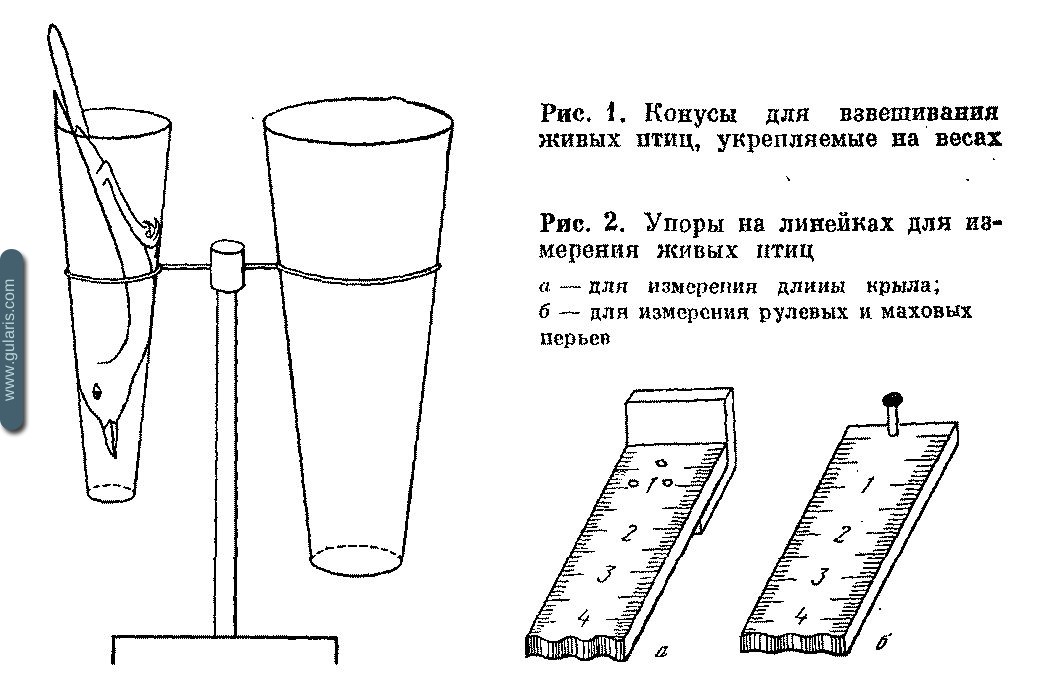
Степень искажения результатов, получаемых кольцеванием, в значительной мере зависит от соблюдения этих простых приемов. Обрабатывать птиц нужно за столом, в помещении. Для работы необходимо следующее оборудование.
1. Быстровзвешивающие весы с ценой деления 0,05—0,1 г. Наиболее пригодны весы ВЛТК-500, на которых установлен держатель для конуса — фиксатора птицы. На Биологической станции ЗИН АН СССР применяют фиксатор, имеющий форму усеченного конуса, склеенный из рентгеновской пленки. Птицу опускают в конус с широкого конца вниз головой, и она заклинивается собственным весом. Конус вставляют в кольцо держателя, укрепленное на весах , и производят взвешивание. Вынимают птицу, перевернув конус широким концом вниз. Птица выскальзывает в руку. Удобно иметь на весах обойму из двух-трех конусов разного размера, соответственно вариации размеров птиц. Следует выдерживать один из стандартов — либо всегда взвешивать птиц с надетым кольцом, либо до того, как оно надето. Средний вес колец каждой серии должен быть известен. При взвешивании окольцованных птиц его вычитают из веса. Последнее лучше осуществлять изменением нулевой настройки шкалы весов, чем арифметической операцией с весом каждой птицы.
2. Линейка для измерения длины крыла — тонкая металлическая или пластмассовая, снабженная упором по линии нулевого деления для кистевого сгиба крыла. Упор делается из металлической или пластмассовой пластинки и прикрепляется заклепками. Измерять крыло штангенциркулем нельзя, так как результат варьирует от степени прогиба крыла.
3. Линейка с перпендикулярным круглым стержнем — упором на уровне нулевого деления — для измерения длины хвоста и отдельных маховых перьев.
4. Короткая линейка для измерения клоакального выступа.
5. Штангенциркуль — для иных промеров.
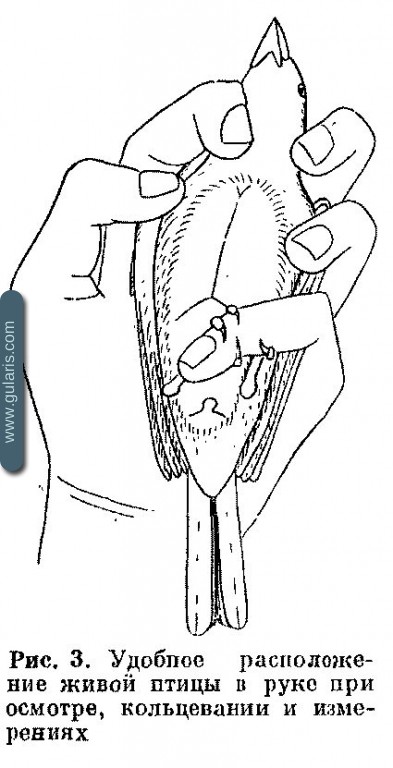
При осмотре птицу держат одной рукой, брюшком вверх, придерживая концами пальцев крылья и голову (рис. 3). В такой позе рассматривают крыло, измеряют его, раскрыв, описывают линьку маховых, надевают кольцо. Для дальнейшего осмотра начинают раздувать перья. Дуть следует не сильно, под острым углом сзади, под перо, одновременно поворачивая птицу так, чтобы струя воздуха последовательно обнажала аптерии. В это время рассматривают клоакальный выступ, если необходимо, измеряют его, рассматривают наседное пятно, оценивают жировые резервы, описывают линьку перьев тела. При этой же процедуре можно отмечать или собирать внешних паразитов. После этого птицу взвешивают и отпускают. Осмотр ускоряется, а ошибки уменьшаются, если запись ведет другой человек под диктовку. При отсутствии у птицы линьки весь осмотр у тренированного человека занимает 15—30 сек., а при линьке — 1—3 мин.
Длина крыла — основной промер, совершенно обязательный для многих целей. Промеры других частей тела производятся, если они необходимы для определения вида, пола или возраста.
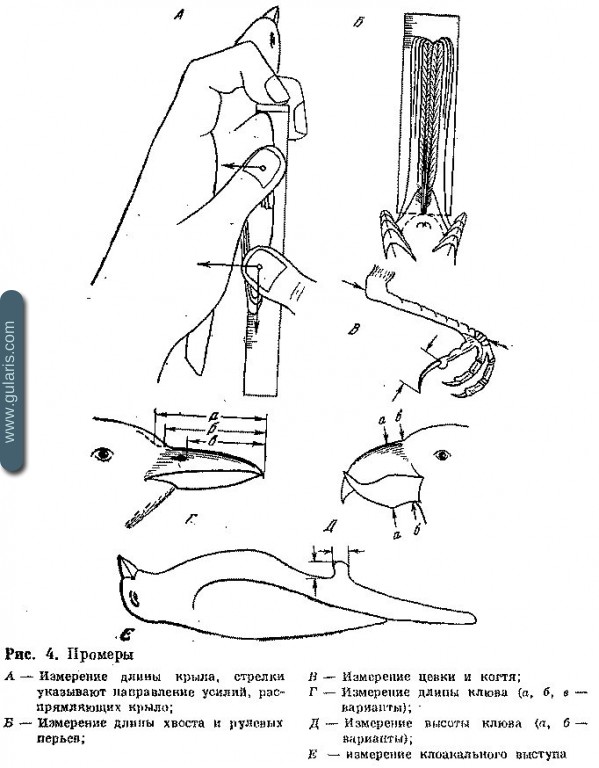
Не следует увлекаться традиционными для коллекционных материалов промерами хвоста, длины тела, размаха крыльев, длины клюва, голени, цевки, пальца, когтя, отдельных маховых и т. д., если не поставлено специальной цели. Длина крыла — это расстояние между кистевым суставом и вершиной самого длинного махового, измеренное при максимально сложенном и прижатом к телу крыле. Существуют три способа измерения длины крыла.
а. Минимальная хорда крыла. Карпальный сустав упирается в нулевой упор, крыло располагается внешним краем вдоль края линейки, касаясь вершиной соответствующего деления. Это устаревший метод, так как он дает большую вариацию. Промеры, полученные этим методом, содержатся в старых определителях.
б. Прижатое крыло — действия те же, но в конце крыло большим пальцем прижимается к линейке, вследствие чего длина увеличивается по сравнению с предыдущим методом на 0,5—3 мм (до 2,5%), а стандартность измерения повышается вследствие устранения прогиба крыла. Измерения этим методом приведены в большинстве современных работ.
в. Прижатое и выпрямленное крыло — действия те же, что и в первом случае. Крыло прижимается большим пальцем одной руки в области крылышка к линейке. Большим пальцем другой руки дистальная часть маховых прижимается к линейке и поворотом этого пальца изогнутый конец крыла выпрямляется вдоль делений, затем движением пальца по ходу нарастания делений вытягивается; одновременно первый большой палец распрямляет вдоль делений среднюю часть крыла и не позволяет карпальному суставу отходить от упора. Получаемый промер дает максимальное и наименее варьирующее значение. Он дает превышение на 0,5—0,7 мм (0,5—6%) в сравнении с первым методом и 0,5—6 мм (0,5—5%) по сравнению со вторым. Этот метод рекомендован в большинстве современных руководств по кольцеванию и прижизненному анализу птиц; им выполнены измерения в настоящем справочнике.
Измерения иных частей тела — длины и высоты клюва, длины цевки и когтя, а также размеров клоакального выступа показаны на рис. 4. При измерении длины клюва обычно указывается, от какого места при его основании начинали измерение (от начала черепа, от границы оперения лба, от ноздрей, границы щетинок и т. д.). Во всех измерениях дается не периметр, а кратчайшее расстояние по хорде. Клоакальный выступ нельзя измерять штангенциркулем или измерителем. Лучше всего пользоваться небольшой линейкой, приложенной сбоку дважды — сначала впереди выступа, слабо уперев нулевое деление в ткани брюшка, а затем повернув линейку делениями к брюшку, сделать второй промер.
Методы регистрации линьки, жировых отложений, стадии наседного пятна и других признаков описываются в следующих разделах.
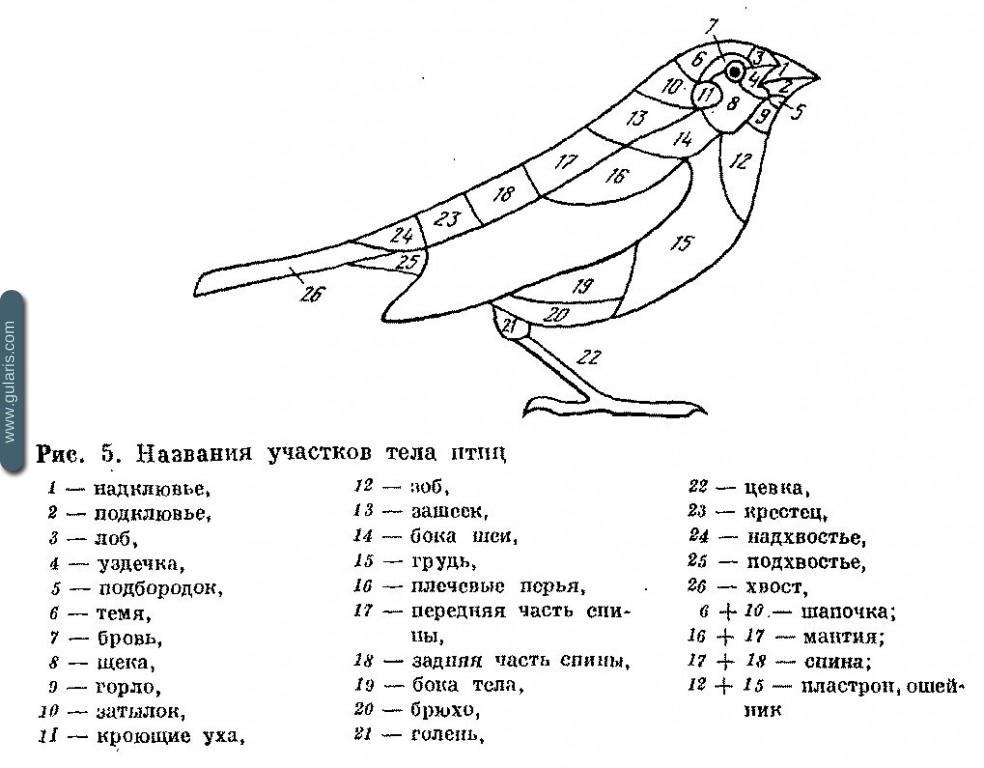
Названия отдельных участков тела
Упоминаемые в повидовой части названия отдельных участков тела можно найти, воспользовавшись рис. 5. Помимо них, в тексте употребляются названия для характерных рисунков: шапочка, пластрон, ошейник, мантия, зеркальце крыла. Наименование перьев в крыле можно установить, пользуясь рис. 6. В тексте часть этих названий дана сокращенно (см. список сокращений).
Нумерация маховых. При работе с разными отрядами птиц вследствие разного числа первостепенных и второстепенных маховых рациональная нумерация осуществляется раздельно для первостепенных и второстепенных, от границы между ними, центробежно. Однако при работе только с воробьиными птицами преимущества такой нумерации оказываются несравненно ниже ее недостатков, и в отношении живых воробьиных основные руководства по кольцеванию и прижизненному анализу рекомендуют более удобную нумерацию, которая осуществляется сквозным порядком, от дистального конца крыла к телу. Этот способ намного удобнее, снижает возможность ошибки (особенно вследствие нечеткой границы между первостепенными и второстепенными, характерной для воробьиных) и не приводит к разночтениям, так как полное число первостепенных маховых у воробьиных птиц нашей фауны одно и то же: десять. Первое, дистальное, первостепенное маховое может укорачиваться, сходя иногда почти на нет. Это имеет место у всех представителей семейств Alaudidae, Hiriindinidae, Motacillidae, Bombycillidae, Fringillidae, Emberizidae.
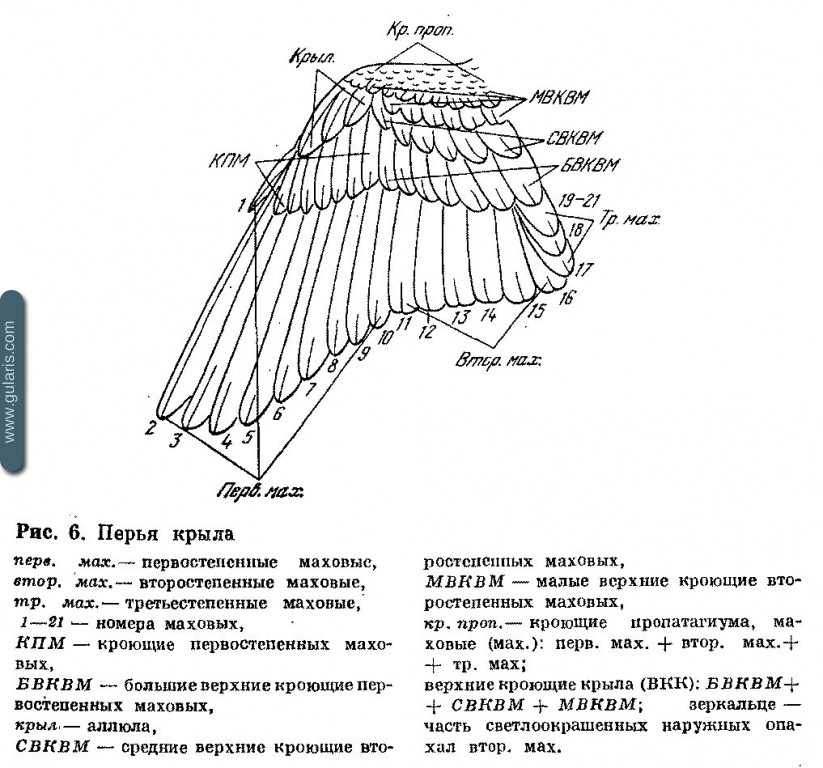
В других семействах первое маховое легко обнаруживаемо, но все же укорочено. Однообразность счета достигается просто: счет начинается с первого полноценно развитого махового, которое всегда второе, поэтому ему независимо от наличия предыдущего пера присваивается второй номер. Далее следует номер третий и т. д. Второстепенных маховых шесть. За ними следуют три третьестепенных маховых. Они лежат одно над другим и опахалами заходят на тело птицы. Четыре третьестепенных обнаружено у Alaudidae и некоторых Corvidae: Perisoreus infaustus, Garrulus glandarius, Pica pica, Nucifraga caryocatactes и Corvus monedula. Остальные Corvidae имеют пять третьестепенных маховых. У Oriolus oriolus пять или четыре третьестепенных (Svensson, 1970).
Так как счет маховых ведется по направлению к телу, вариация количества третьестепенных не влияет на идентичность нумерации предыдущих перьев. В настоящем справочнике нумерация перьев приводится по описанному выше принципу, причем иногда указывается просто порядковый номер пера по сквозной нумерации (например, двенадцатое перо есть второе второстепенное), а иногда раздельно при первостепенных, второстепенных и третьестепенных (например, пятое второстепенное, второе третьестепенное).
Обозначения возрастов
Принятая EURING система обозначения возрастов птиц при кольцевании (гнездовой птенец, слеток, полностью выросший, первогодок, второго док), к сожалению, не может быть полностью использована при описании признаков возраста, так как последние чаще всего базируются на нарядах. Поэтому границы возрастов приходится коррелировать с линьками, А так как не все виды имеют одинаковые сроки линьки, соответственно этим срокам длительность пребывания птицы в определенном возрастном наряде различна. В настоящем справочнике приняты следующая система обозначения возрастов и следующие границы возрастов:
pull — гнездовой птенец до вылета из гнезда; наряд ювенильный, формирующийся.
juv — от вылета из гнезда до завершения первой, постювенильной линьки. У видов с полной постювенильной линькой верхняя граница возраста проходит по той стадии линьки, на которой утрачивается возможность отличать по внешнему виду первогодков от взрослых птиц. Если постювенильная линька у вида происходит зимой, обозначение птиц в этом возрасте, соответственно, продолжается до зимней линьки.
sad — от окончания постювенилыюй линьки до первой послебрачной линьки.
ad — взрослая птица в дефинитивном оперении, начиная с первой послебрачной линьки.
Обозначения сезонов
Вследствие постювенильной, предбрачной и послебрачной линек, обнашивания оперения и существования дополнительных сезонных признаков пола и возраста для большинства видов приводимые в справочнике критерии пола и возраста даны раздельно по сезонам. При этом фактически под тем или иным сезоном подразумевается не определенный календарный сезон, а определенное сезонное физиологическое состояние птиц, сроки которого варьируют у разных видов в пределах 1—3 месяцев. Применены следующие названия сезонов (которые фактически сокращенно обозначают сезонные физиологические состояния):
Весна — от окончания предбрачной линьки (если она есть у этого вида), развития половой активности и начала весенней миграции до окончания весенней миграции и начала размножения.
Лето — от прилета и начала размножения до окончания размножения и начала летней линьки.
Осень — от окончания линьки и (или) начала осенней миграции до завершения ее.
Зима — от завершения осенней миграции до начала весенних процессов. В случаях, сильно уклоняющихся от этого стандарта, сезон поясняется более подробно.
ПРИЗНАКИ ПОЛА И ВОЗРАСТА
Мы научились различать самцов, самок, молодых и взрослых не у всех видов птиц. Однако можно утверждать, что возможно опознавать пол и возраст у большинства, если не у всех видов. Самим птицам для избегания ошибок в поведении при стайных, иерархических, территориальных и брачных контактах необходимо, по крайней мере, в некоторых ситуациях опознавать пол и возраст партнеров. И действительно, мы очень часто убеждаемся, что в конце концов удается найти нужные признаки у вида, в отношении которого долгое время различение пола или возраста казалось невозможным.
Разделим признаки на (1) внешние, видимые морфологические различия, используемые для опознания и самими птицами и исследователем — они наиболее удобны и хорошо известны; (2) обнаруживаемые исследователем, но, видимо, не используемые самими птицами признаки — такие, как пневматизация черепа, наличие наседного пятна или форма клоакального выступа и (3) поведенческие сигналы (позы, движения, звуки), доступные для опознания и птицам и исследователю, но только при наблюдении за естественным поведением.
Признаки последней группы непригодны при кольцевании и работе с тушками. Признаки второй группы пригодны при кольцевании, но непригодны для коллекционной работы. В первую группу признаков входят как сохраняющиеся в тушках признаки, так и исчезающие (например, цвет глаз).
Кольцующий живую птицу исследователь имеет больше возможностей для определения пола и возраста по сравнению с исследователем, обрабатывающим коллекционный материал. В летний период пол полевого воробья при наблюдении за свободноживущими птицами определяется за несколько минут по характерным позам самца и самки; у пойманного воробья он сразу определяется по наседному пятну и клоакальному выступу; но у тушек пол этого вида неопределим.
В этом разделе мы, во-первых, приводим признаки пола и возраста, применимые к многим видам. Описываются признаки, их вариации и ограничения применения. В диагнозах для разных видов они каждый раз не описываются, а приводятся только ссылки на настоящий раздел. Во-вторых, поскольку работа по обнаружению признаков пола и возраста у всех видов еще не завершена, мы специально останавливаемся на оказавшихся эффективными приемах поиска новых признаков, особенно у живых птиц.
Признаки пола
Некоторые признаки пола проявляются очень рано, и в таких случаях опознание пола возможно уже у птенцов или слетков. Таковы цвет определенных частей оперения у некоторых Paridae. У Sturnus vulgaris пол определяется по цвету радужины через три недели после вылета; у некоторых видов появляются отдельные перья самцовой окраски (Fringilla coelebs, Spinus spinus).
Другие признаки связаны с определенным сезонным физиологическим состоянием и проявляются только во время этого состояния. Таковы изменение формы клоакального выступа, развитие наседного пятна, изменение цвета клюва. Третья группа признаков ежегодно появляется при одной смене наряда и исчезает при следующей смене наряда.
Наконец, многие признаки, проявившись в определенном возрасте, сохраняются до конца жизни.
Чтобы надежно применить признак для определения пола, мы часто должны предварительно определить возраст птицы, ее сезонное физиологическое состояние и положение относительно цикла смены нарядов.
Размеры
У многих видов самцы и самки различаются по размерам и весу тела. Вес тела одной особи может сильно варьировать в следствие отложения жира, поэтому он, особенно у перелетных птиц, не надежный признак. Но вес птиц определенного уровня жирности, в частности, птиц без видимых жировых отложений,— хороший признак. В повидовых диагнозах в настоящем справочнике он приведен для тех видов, по которым материал достаточен.
Длина крыла — наиболее употребимый размерный признак. У некоторых (очень немногих) видов пределы индивидуальной вариации длины крыла самцов и самок не перекрываются, и тогда это прекрасный признак для определения пола каждой особи.
Чаще пределы вариации длины крыла самцов и самок перекрываются. Тогда мы можем определить пол птицы, если длина ее крыла оказывается вне зоны перекрытия.
Для успеха многих исследований достаточно, чтобы пол был определен у части окольцованных птиц и имелась бы уверенность в том, что в группу птиц с неопределенным полом самцы и самки попадают в равном соотношении. Поэтому указание пола не у всех кольцуемых птиц допустимо. В видовых диагнозах приведены для каждого пола средняя длина крыла и пределы ее вариации. Если число измеренных птиц (которое также приведено) более 100, определение пола птицы с длиной крыла вне зоны перекрытия верно с 99—95%-ной вероятностью. Если число измеренных птиц меньше 100, не следует определять пол по длине крыла, близкой к зоне перекрытия. Возрастные различия в длине крыла, если возраст птиц неопределим, уменьшают надежность признака.
Сезонные изменения длины крыла в популяции обусловлены изменением возрастного состава, избирательной смертностью и постепенным снашиванием концов перьев после линьки. У птиц с длиной крыла 70—90 мм укорочение крыла за счет снашивания может составлять 1—4 мм, причем у молодых птиц с неполной постювенильной линькой снашивание больше, чем у взрослых. Авторы имели по некоторым видам ограниченный материал, поэтому не могли дать полные характеристики крыла для всех видов. Приступающий к кольцеванию орнитолог имеет возможность, пользуясь приведенными выше рекомендациями, составить для отлавливаемых в достаточном количестве видов полные характеристики вариации длины крыла, по которым не только будет определять пол в дальнейшем, но и установит пол уже окольцованных птиц, если при кольцевании он записывал длину крыла. Вторжение перелетных птиц других популяций того же вида, имеющих более длинное крыло, на территорию, заселенную короткокрылой формой (или наоборот), может приводить к ошибкам в определении пола.
Чисто методические трудности, наконец, затрудняют применение длины крыла как критерия пола: приводимые в литературе данные о пределах вариации длины крыла могли быть получены при другой технике измерений, на птицах иной популяции, иного…
Промеры других частей тела в некоторых случаях служат для определения пола. Пол у некоторых видов хорошо определяется по длине хвоста или его частей (например, у Hirundo rustica, Н. smithii, Terpsiphone paradisi).
Различия в окраске оперения
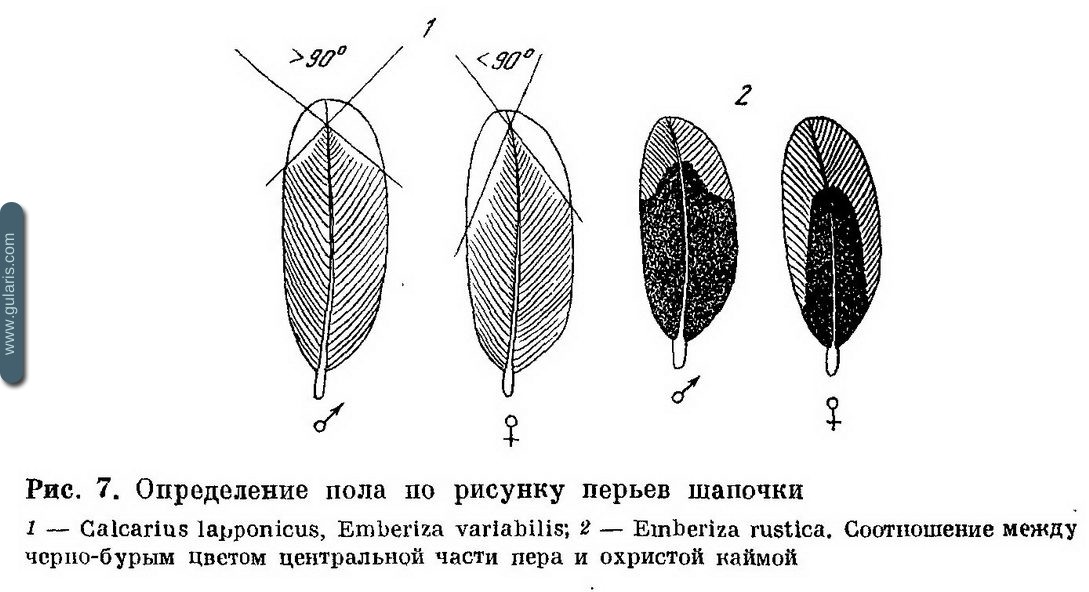
Когда различия в рисунке или окраске сразу видны, они не описываются подробно, так как входят в любые определители. Но многие диагнозы построены на малозаметных различиях в рисунке или па слабых различиях в цвете или оттенке. Все же они таковы, что человек с нормальным зрением может научиться определять птиц по приводимым признакам. Для этого сначала лучше определять не одну птицу, а осматривать серию и, пользуясь указаниями диагноза, обнаружить вариацию по указанному признаку, а затем опознать ту границу, по которой разделяются птицы разного пола. При обучении не следует обращать внимания на другие признаки, по которым в серии намечаются различия.
Почти наверняка авторы проверяли эти признаки, но отвергли их как не связанные с полом. Нужно сконцентрировать все внимание на указанном признаке. Определение пола в некоторых случаях требует очень высокой квалификации и терпения. Но, освоив признак, орнитолог без затруднений пользуется им. На рис. 7 приведены примеры малозаметных, но надежных критериев пола, основанных на различиях в окраске отдельных перьев.
Цвет радужины глаза
Это прекрасный признак для некоторых видов. Так как он виден только на живых птицах, авторы приводят его для тех видов, которые изучены ими в живом виде. Приступающий к кольцеванию орнитолог должен в отношении своих видов проверить, составлены ли диагнозы с использованием живых птиц или только по тушкам. Если живой материал не изучен, следует обратить внимание на вариации цвета радужины с целью выяснить их диагностическую ценность.
Цвет клюва
У некоторых видов полы во все сезоны различимы по окраске клюва. У других видов окраска клюва одного из полов изменяется на время половой активности, и в этот период, т. е. во время весенней миграции и размножения, может служить хорошим диагностическим признаком для определения пола. В повидовых диагнозах этот признак использован нами редко, так как, во-первых, он плохо сохраняется у тушек, во-вторых, часто у того же вида есть и другие признаки. Приступающий к кольцеванию орнитолог должен проверить, нет ли вариации в цвете клюва у отлавливаемого им вида, и если она есть, установить возможность использования этого признака для определения пола.
Форма клоакального выступа
Практически у всех воробьиных развитие половой активности сопровождается изменением формы клоакального выступа. Изменения различны у самцов и самок (рис. 8), и к концу весенней миграции различия между самцами и самками становятся столь отчетливыми, что позволяют вплоть до конца размножения определять пол почти у всех видов. Попытки использовать этот признак во время линьки, осенью, зимой и ранней весной необоснованны. В неактивный период клоакальный выступ у обоих полов невелик.
С развитием половой активности у самцов объем клоакального выступа увеличивается в 10—70 раз. Выступ булавовидно вздувается; передняя граница его с брюшком становится резкой и часто образует складку; продольная ось клоакального отверстия смещается вперед. У самок изменения вначале менее заметные. К началу спаривания и во время откладки яиц высота клоакального выступа изменяется мало, но его диаметр увеличивается, отверстие, расширяясь, приобретает овальную форму; продольная ось отверстия смещается назад. Птицы способны при раздражении втягивать клоакальный выступ в тело, поэтому при осмотре его не следует трогать.
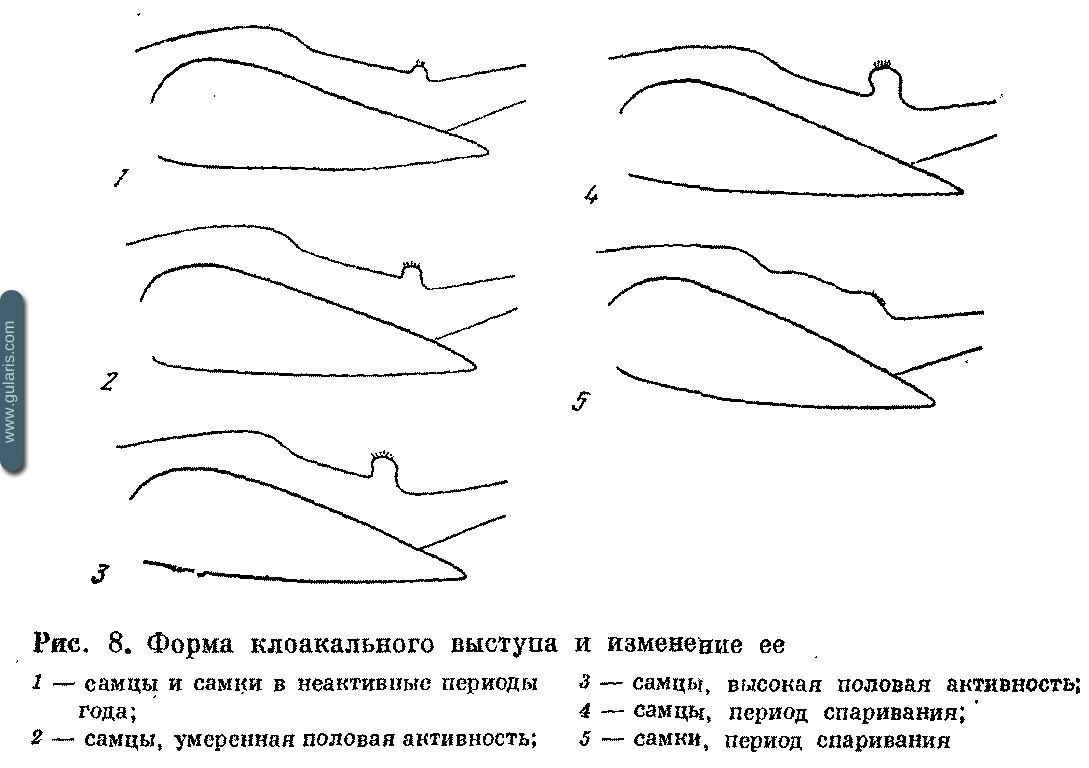
Существует видовая вариация в степени проявления половых различий в форме и размерах выступа, но неизвестны случаи, чтобы выступ самцового типа развивался у самок и наоборот. Поэтому это самый универсальный признак пола во время размножения. Он слабо выражен у Hippolais icterina, Oriolus oriolus, Sturnus vulgaris.
Наседное пятно
На время размножения у самок развиваются наседные пятна. Наседное пятно охватывает всю вентральную поверхность — от начала грудины до клоаки, и хорошо обнаруживается при раздувании перьев. В своем развитии наседное пятно проходит ряд морфологических изменений, по которым могут быть выделены отдельные стадии, удобные для регистрации (подробно о наседном пятне см. Bailey, 1954; Petersen, 1955; Люлеева, 1965, 1967; Ефремов, Паевский, 1973). Сопоставление этих стадий с положением птицы относительно цикла размножения показало столь отчетливую связь, что открывается возможность по стадии наседного пятна пойманной самки устанавливать ее положение в гнездовом цикле.
Как правило, используются четыре основных стадии, которые можно обнаружить во всех случаях. При более подробных исследованиях вводится несколько промежуточных стадий.
I стадия — «очищение от перьев». Кожа гладкая, тонкая, глянцевая. Происходит очищение поверхности будущего наседного пятна от перьев. Первая стадия, как правило, у большинства видов начинается во время гнездостроения и длится несколько дней, завершаясь к откладке первого яйца.
II стадия — «васкуляризация». На поверхности пятна появляется густая сеть видимых сквозь кожу кровеносных сосудов. Кожа остается гладкой, но к концу стадии мутнеет и утолщается. Вторая стадия длится несколько дней, завершаясь к началу насиживания.
III стадия — «отек». Развивается отечность кожи вследствие увеличения содержания в ней лимфы. Кожа становится толстой, мутной, сосуды перестают просвечивать, поверхность кожи гладкая. По внешнему виду наседное пятно напоминает пузырь при ожоге. Третья стадия начинается одновременно с началом насиживания и продолжается до достижения птенцами возраста 5—7 дней. Максимально развита в момент вылупления, после чего отечность уменьшается, кожа становится дряблой и образует складки. Последняя часть процесса, связанная с согреванием птенцов, иногда выделяется в качестве дополнительной стадии.
IV стадия — «восстановление». Отечность пропадает, кожа становится сухой, сморщивается, шелушится, вновь начинают просвечивать кровеносные сосуды. Четвертая стадия длится со второй половины периода выкармливания птенцов в гнезде до послебрачной линьки. К вылету молодых кожа уже сухая, плотная, шелушащаяся. В четвертой стадии можно выделять две промежуточные стадии. Размножавшиеся самки узнаются до линьки по не заросшему перьями участку, на котором было пятно.
Если размножение повторяется, вновь образуется отек и потом восстановление. Таких самок в стадии отека можно узнать по остаткам шелушения кожи, которого при первом отеке не бывает.
Если гнездование прервано, наседное пятно сморщивается стремительно, образуя сухие коросты. Таких самок также можно отличить от окончивших размножение удачно. Не следует смешивать наседное пятно с незаросшим голым брюшком молодых птиц. Различение пятна и незаросшего брюшка молодых достигается после некоторой практики на ясных в отношении возраста видах. Есть указания, что в некоторых случаях у части воробьиных птиц развитие наседного пятна может начинаться до начала размножения. О такой возможности следует помнить при работе с малоизученными видами или в специфических условиях.
Применимость наседного пятна как критерия пола ограничена. Во-первых, не участвующие в размножении самки наседных пятен не имеют. Во-вторых, у некоторых видов наседное пятно образуется и у самцов — например, у большинства видов Sylvia, у Nucifraga caryocatactes, Passer hispaniolensis и, по-видимому, у других видов. У всех этих видов самцы участвуют в насиживании. Но у других видов, несмотря на участие в насиживании, самцы наседных пятен не образуют. Наседное пятно можно применять как признак пола для видов, у которых насиживают только самки. Если же оба пола окрашены сходно и участие полов в насиживании неизучено, этот признак требует обязательной проверки. Не следует доверять беглым указаниям в фаунистических работах о поведении полов при насиживании, так как они могут быть ошибочными.
При обнаружении наседного пятна у сходно окрашенных полов нужно сопоставить наличие наседного пятна у птицы с формой клоакального выступа: у самцов, имеющих наседное пятно, выступ остается по форме типичным для самца.
Признаки возраста
Некоторые признаки возраста одинаково применимы от вылета из гнезда до первой послебрачной линьки; другие ограничены более короткими периодами или, сначала проявляясь четко, постепенно угасают. Очень многие из этих признаков связаны с особенностями ювенильного оперения, неполнотой первой постювенильной линьки и с различиями в форме и окраске оперения двух смежных нарядов.
Молодые птицы (Juvenilis)
Только что покинувшие гнездо птенцы обычно без труда отличаются от взрослых по внешнему облику. Оперение у них мягкое, более рассученное и пушистое, чем у взрослых. На теле долго сохраняются голые участки — на груди, на брюхе, в подмышечной части и на нижней стороне крыла. Хвосты могут быть короткими, в стадии дорастания. Маховые часто уже, заостренней и короче, чем у взрослых; редуцированные первостепенные маховые, напротив часто длиннее и шире, чем у взрослых. Особенно удобно для диагностики возраста первое маховое. У тех видов, для которых в дефинитивном оперении характерна редукция первого махового, в ювенильном оперении это маховое длиннее, шире и закругленнее, чем в следующих нарядах (рис. 9). Если постювенильная линька полная, этот признак позволяет отличать juv, с одной стороны, и sad и ad — с другой. Если постювенильная линька не затрагивает маховых, этот признак отличает juv и sad от ad. Первое маховое во время линьки сменяется последним, поэтому определение возможно до конца линьки. Так как у птенцов маховые и рулевые растут все одновременно, остатки чехлов обнаруживаются сразу в основании всех полетных перьев и не могут быть спутаны с линькой взрослых, поскольку у последних перья сменяются неодновременно: первостепенные маховые в дистальном направлении, второстепенные маховые — центростремительно, а рулевые — центробежно 1. Глаза у некоторых видов имеют в ювенильный период иную окраску. В углах клюва есть вздувшиеся валики, обычно желтого цвета (не смешивать с вздутыми валиками у кормящих взрослых; у последних они оранжевые с неровной, морщинистой поверхностью).
Молодые сохраняют некоторое время особый цвет полости клюва и пятна на языке (см. Locustella). Ноги обычно мягче и светлее. Иногда есть различия в цвете клюва и в его форме: у взрослых конец надклювья выходит за кончик подклювья и несколько загнут книзу; у молодых концы надклювья и подклювья совпадают.
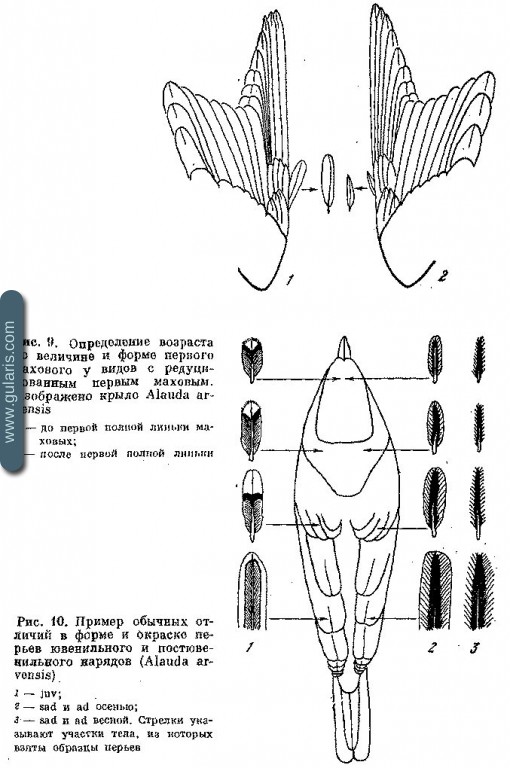
Большинство перечисленных ювенильных признаков постепенно исчезает в течение нескольких первых недель жизни. Особенности окраски птенцового оперения сохраняются вплоть до второй половины постювенильной линьки. Молодым птицам многих видов свойственна особая окраска, которая имеет тенденцию быть более незаметной за счет избегания ярких цветов и образования рисунков чаще всего в форме штрихов, мелких пятен, струйчатости, чешуйчатости (рис. 10). Светлый низ тела часто имеет сероватый или желтоватый оттенки. Если в окраске оперения молодых есть особенности, отличающие их от взрослых, то их удается узнавать почти до самого окончания постювенильной линьки, находя среди нового оперения отдельные перья ювенильного наряда (см. рис. 10).
Если даже молодые птицы окрашены так же, как взрослые, они при некотором навыке отличаются от них во второй половине лета по степени обношенности оперения, которая больше у взрослых птиц, носивших наряд много месяцев, по сравнению с молодыми, носящими его первый или второй месяц.
Летние признаки взрослых птиц
При кольцевании различение возрастов облегчается тем, что в одно и то же время встречается вместе ограниченное число нарядов. Так в течение второй половины лета у большинства воробьиных приходится отличать только молодых рождения этого года от взрослых, уже гнездившихся птиц. В течение всего периода до линьки и в начале линьки взрослые птицы легко опознаются по наличию у самок наседного пятна и по увеличенному клоакальному выступу у самцов. В конце первой трети линьки и позднее эти признаки исчезают, но определение возраста возможно по другим признакам, связанным с различиями в нарядах и с особенностями линьки разных возрастных групп.
Линька
У большинства воробьиных постювенильная и послебрачная линьки происходят во второй половине лета и осенью и лишь у части видов зимой. В большинстве случаев постювенильная линька бывает неполной, т. е. часть оперения не сменяется. Это различие в полноте постювенилыюй и послебрачной линек позволяет узнавать молодых птиц в течение всего первого года жизни по отличию сохранившихся ювенильных перьев от соответствующих перьев взрослого наряда. У некоторых групп постювенильная линька полная, например в семействах Alaudidae, Sturnidae, Ploceidae, а также у некоторых видов из других семейств (Panurus biarmicus, Aegithalos caudatus, Emberiza calandra и др.). У этих видов определение возраста по оперению становится невозможным с конца линьки. Следующие замечания о характере линек разных групп воробьиных (в пределах обитающих в СССР видов) могут быть полезны при изучении и использовании признаков пола и возраста по оперению:
Послебрачная линька. Один раз в год, полная, в конце лета — осенью: Alaudidae, Corvidae, Motacillidae, часть Laniidae, Sturnidae, Bombycillidae, Cinclidae, Troglodytidae, Prunellidae, многие Sylviidae, Agrobates galactotes, Regulidae, Muscicapa cyanomelana, M. albicollis, M. mugimaki, M. hypoleuca, M. parva, большинство Turdidae, Paridae, Sittidae, Certhia, Paradoxornithidae, Tichodroma, Ploceidae, Fringillidae, кроме Carpodacus erythrinus, Emberizidae. Один раз в год, полная, после осенней миграции: большинство Hirundinidae, часть Laniidae, Oriolus chinensis, Carpodacus erythrinus; самцы Oriolus oriolus. Полная, но разделена на две части, разделенные осенней миграцией: некоторые Hirundinidae, часть Laniidae, часть Sylviidae, Muscicapa striata, М. griseisticta, M. sibirica, M. latirostris, самки Oriolus oriolus 1.
1 Исключение составляет Muscicapa striata, линяющая по другой схеме.
Постювенильная линька. Один раз в год, полная, в конце лета и осенью: Alaudidae, Sturnidae, Lrosphena squameiceps, Paradoxornithidae, Certhia, Ploceidae, Emberiza calandra. Один раз в год, полная, после осенней миграции: большинство. Hirundinidae. Один раз в год, частичная, в конце лета — осенью: Corvidae, Motacillidae, некоторые Laniidae, Bombycillidae, Cinclidae, Troglodytidae, Prunellidae, часть Sylviidae, Regulidae, Muscicapa parva, большинство Turdidae, Paridae 2, Sittidae, Tichodroma, 1 Маховые и рулевые, исключая центральную пару, сменяются на зимовках. 2 Включает факультативную линьку рулевых у некоторых видов. Fringillidae, кроме Carpodacus erythrinus, Emberizidae. Один раз в год, частичная, после осенней миграции: Oriolus chinensis, Carpodacus erythrinus. Разделена на две части, прерванные осенней миграцией, в сумме полная: Oriolus oriolus, Muscicapa striata, M. griseisticta, M. sibirica. Разделена на две части, прерванные осенней миграцией, в сумме частичная: некоторые Laniidae, Muscicapa hypoleuca, M. latirostris, M. albicollis, М. mugimaki, М. cyanomelana. Варьирует индивидуально по признаку долей оперения, сменяемых до и после осенней миграции, в сумме частичная: некоторые Laniidae.
Предбрачная линька *. У взрослых полная: часть Sylviidae, Muscicapa cyanomelana, M. albicollis, M. mugimaki. У взрослых частичная: Motacillidae 2, часть Laniidae (иногда почти полная), часть Sylviidae, Muscicapa hypoleuca, M. parva, часть Turdidae, большинство Emberizidae 3. У молодых полная: часть Sylviidae. У молодых частичная: Motacillidae (кроме контурных сменяются отдельные маховые и рулевые), часть Laniidae (иногда почти полная), часть Turdidae, Remiz, большинство Emberizidae. Характер линек не изучен. Zosteropidae (видимо, две линьки), Campephagidae, Pycnonotidae, Muscicapa narcissina, M. ruficauda, Terpsiphone paradisi.
Неполнота постювенильной линьки у близких видов, разных подвидов и популяций одного вида и даже особей из ранних и поздних выводков обнаруживает зависимость от географического положения. В целом южные и юго-западные популяции имеют больше времени для линьки, и линька у них более полная по сравнению с северными и северо-восточными популяциями. Поздно родившиеся особи могут линять менее полно, чем рано родившиеся. Наконец, в некоторых случаях часть линьки или вся линька может переноситься на период после осенней миграции. Все эти особенности, насколько они известны, указаны в повидовых описаниях. Но о них следует помнить при работе с малоизученными видами или на новых территориях. Рассмотрим подробнее наиболее частые пути определения возраста по различиям в постювенильных линьках.
Смена маховых и рулевых перьев у видов с неполной постювенильной линькой происходит только у птиц старше одного года. У таких видов в период линьки особь, сменяющая маховые и рулевые — взрослая. Чтобы избежать ошибок, следует проверять симметричность линьки на обоих крыльях; хвост менее надежен, так как рулевые могут быть случайно утрачены.
1 Смена оперения (полная или частичная) в течение зимы или весны называется предбрачной линькой, если в конце лета — осенью или в начале зимы ей предшествовали нослебрачная или постювенильная линьки. Поэтому у Motacillidae весной контраст в цвете и обношенности между отдельными перьями — не показатель возраста. Е. melanocephala имеет еще одну частичную линьку после размножения; сменяются третьестепенные маховые.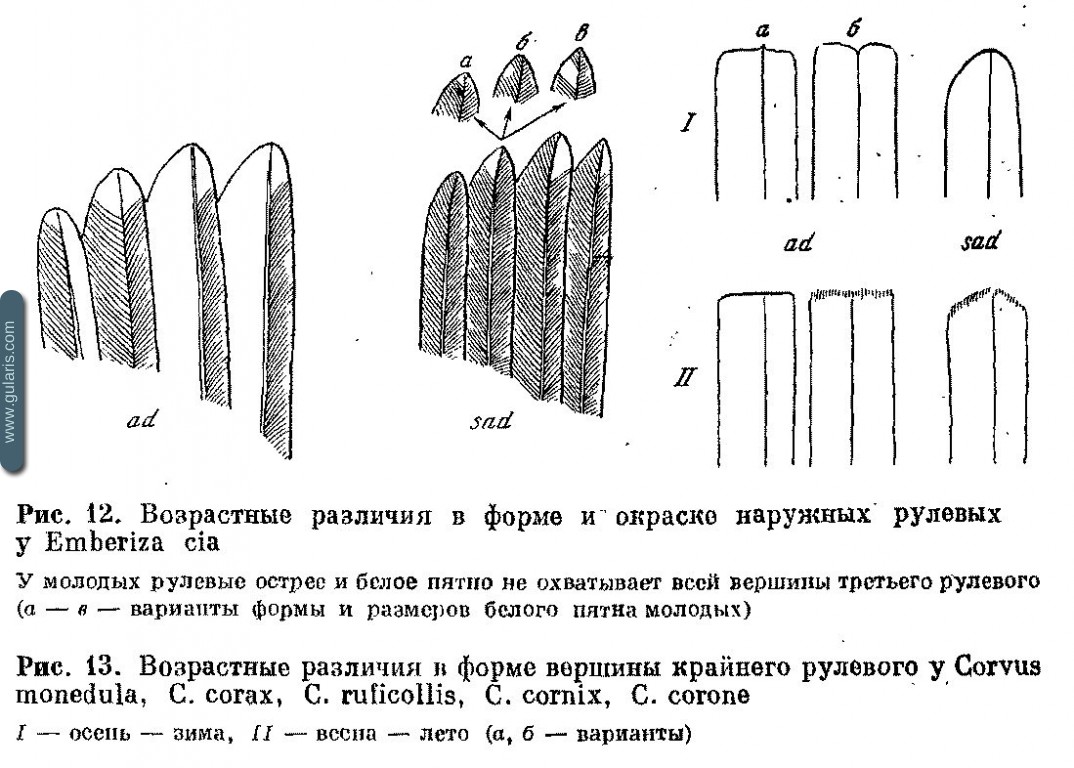
У большинства воробьиных 6-е второстепенное маховое (нумерация в направлении к телу, 16-е перо от вершины крыла) сменяется последним и растет медленнее других. В конце линьки остатки чехла на этом пере — признак взрослой птицы; несколько ранее такой же указатель — чехлы на других маховых. Три третьестепенных маховых у воробьиных (17, 18 и 19-е при нумерации по направлению к телу) часто не охватываются постювенильной линькой и могут быть узнаны как ювенильные перья в течение зимы, весны и лета. Они будут рассмотрены подробнее вместе с кроющими крыла. Форма рулевых перьев у многих видов различна в ювенильном и взрослом наряде. Соответственно они могут служить критерием возраста до постювенильной линьки, если она полная и до первой послебрачной, если она частичная. Если рулевые в двух нарядах различаются, то, как правило, ювенильные рулевые (особенно центральная пара) уже, заостреннее, или конец внутреннего опахала имеет более острый угол к стержню, чем у взрослых птиц.
У молодых птиц иногда центральная пара рулевых светлее и менее устойчива к обнашиванию, в результате чего во второй половине первого года жизни более обношена, чем у взрослых. Признак этот следует применять осторожно, так как не у всех видов он отражает возраст, а у тех видов, у которых он его отражает, эта А — полностью непневматизированный череп, D — полностью пневматизированный. Справа внизу — место образования пробора при осмотре живой птицы пара легко утрачивается, после чего заменяется взрослыми перьями.
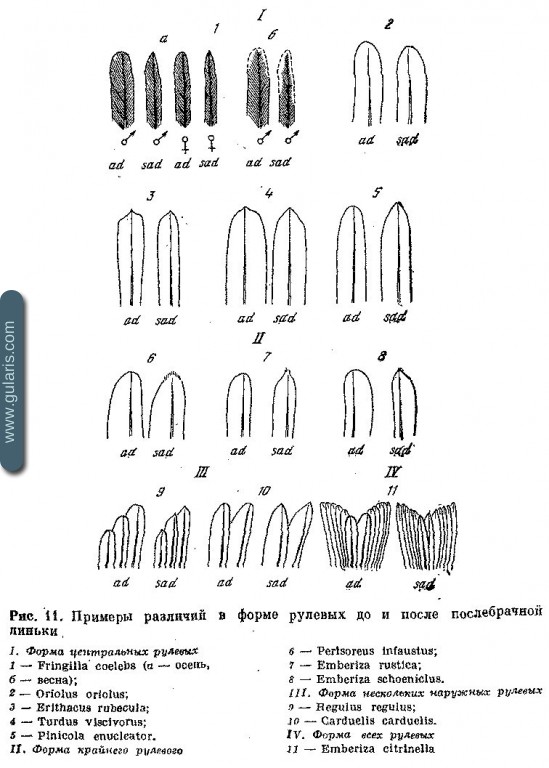
На рис. 11 приведены примеры различий в форме рулевых у нескольких видов. Признак часто встречается у Turdidae, Fringillidae, Emberizidae. Иногда возраст определяется по цвету каемок рулевых (см., например, Cannabina cannabina и рис. 12).
У видов с длинными крайними рулевыми нужно обращать внимание на их обношенность и длину относительно остальных перьев (у молодых обычно короче); у juv Aegithalos caudatus короче центральные рулевые. У некоторых Corvidae ювенильные перья хвоста округлые, а во взрослом наряде тупые (рис. 13).
Неполная смена кроющих крыла при постювенильной линьке позволяет определять возраст у многих видов. Верхние кроющие крыла и третьестепенные маховые в ювенильном и взрослом нарядах могут отличаться по цвету, форме, деталям рисунка или по изношенности.
Цветовые различия, если они существуют, достаточны для определения возраста. У взрослых птиц летом кроющие более обношены, чем у молодых; осенью, зимой и весной, наоборот, они более обношены у молодых. Обнашивание зависит от цвета перьев. Так, черные перья хуже обнашиваются, чем более светлые, но они постепенно изменяют цвет на буроватый. У видов с рыхлым, пушистым оперением обнашивание почти незаметно.
После окончания линьки обнаруживается контраст в цвете, рисунке или обношенности между перелинявшими и не перелинявшими участками, если линька была неполной у молодых. Этот признак широко использован в данном справочнике; чтобы им пользоваться, нужно научиться замечать контрасты между участками кроющих. Для этого крыло рассматривают в ровном дневном свете, вдали от дающих блики белых или ярко окрашенных предметов, поворачивая его под разными углами к источнику света.
Часть признаков более заметна на максимально сложенном крыле, часть на плавно раскрываемом и закрываемом. Можно перемещать перья кончиком пальца или карандаша. Количество сменяемых перьев в крыле при неполной линьке варьирует по видам, популяциям и индивидуально. Поэтому участки старых и новых перьев могут варьировать по положению и площади. Есть опасность, что в некоторых случаях у молодых вылинивают все кроющие крыла или не вылинивает ни одного. В этих случаях контраст не обнаруживается. Но контраст может быть найден при сравнении кроющих с перьями других частей тела. Промеры длины крыла обычно не дают достоверных различий между взрослыми и молодыми. В некоторых случаях в качестве признаков возраста используются различия в длине отдельных перьев (см. Pica pica, Panurus biarmicus, Locustella, Aegithalos caudatus), частей пера (см. Coccothraustes), размерах цветовых пятен (см. Bombycilla garrulus).
Пневматизация черепа
Молодые птицы родятся с непневматизированным черепом, и процесс пневматизации продолжается в течение первых четырех — семи месяцев жизни. Самая быстрая пневматизация описана у Hippolais icterina — в течение двух месяцев; у некоторых видов она длится почти год. Например, Kemiz pendulinus может приступать к размножению с неполностью пневматизированным черепом. Так или иначе, этот признак всегда позволяет установить возраст в течение второй половины лета и большей части осенней миграции (рис. 14). Непневматизированные участки черепа прозрачные, глянцевые, желто-оранжевого цвета, пневматизированные — матовые, непрозрачные, белого цвета с крупитчатой структурой. Этот признак используется для проверки предположений о других внешних признаках возраста при вскрытии птицы. Но он применим и в отношении живых птиц.
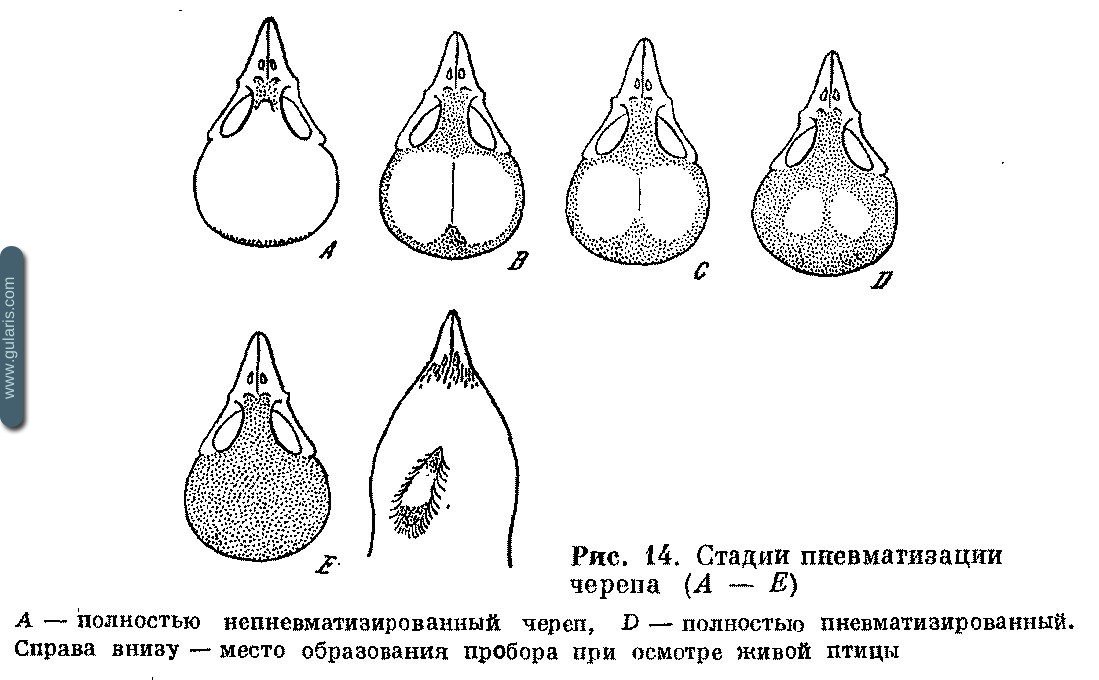
Скальпирование — самый жестокий прием. На теменной части кожи головы делают небольшой надрез, раздвигают края, и через отверстие, передвигая его, находят границу пневматизированной и непневматизированной зоны, если птица молодая. Затем надрез заклеивают хирургическим клеем.
Удаление перьев — небольшой участок кожи на голове освобождают от перьев, смачивают и, передвигая образовавшееся окошко на разные участки черепа, находят просвечивающую сквозь кожу границу пневматизированного и непневматизированного участков. Образование пробора — основной и единственный допустимый к применению в больших масштабах метод. Взяв птицу в левую руку спиной кверху, фиксируют голову двумя пальцами за клюв и щеки, а концом третьего — со стороны затылка. Другой рукой смачивают перья на верхней части головы, после чего раздвигая их влево и вправо от продольной оси головы, образуют пробор, идущий вдоль головы чуть левее или правее продольной оси (см. рис. 14). Кожу в проборе смачивают и через образовавшееся окно, передвигая его, находят границу пневматизированного и непневматизированного участков. Если найти ее не удалось, делают новый пробор в другом месте, пока не найдут границу или не убедятся в ее отсутствии.
П р и м е ч а н и е . Метод может завышать число взрослых птиц, если его применяют в конце миграции, у рано размножающихся, видов, или на зимовках. Если птица ударилась головой в стекло в комнате или отлов производится ловушкой с стеклянным отражателем, то на черепе взрослой птицы образуется отек из лимфы или крови, и она может быть принята за молодую. Желательно накопление данных о сроках завершения пневматизации черепа у разных видов и в разных частях страны.
Предостережение
Прежде чем указывать в записи пол или возраст кольцуемой птицы, нужно твердо убедиться в том, что определение верно. При любых способах отлова птиц какое-то число их погибает, и если на каждой погибшей птице вскрытием проверять пол (по семенникам и яичникам) и возраст (по пневматизации черепа и фабрициевой сумке), то через некоторое время используемый внешний признак будет проверен. При недостаточной уверенности пол и возраст птицы не следует указывать в записи кольцевания. Правильный путь — указать в примечании признак целиком или применив для него сокращенное обозначение. Если обнаруживается набор вариаций разных признаков, следует обозначить все варианты и все их комбинации. В будущем часть их может оказаться признаками пола, возраста, а часть — проявлением иных форм полиморфизма, и все это будет установлено, а пол и возраст ранее окольцованных птиц определен по записям.
Не следует доверять традиционным признакам пола и возраста. В среде птицеловов, охотников и любителей певчих птиц бытуют наряду с верными много «надежных» признаков, не выдерживающих проверки. Приводимые в видовых определителях признаки самцов, как правило, верные, но следует помнить, что очень часто они касаются только старых птиц с полностью развитым нарядом. Если наряд развивается в течение нескольких лет, то только самые старшие самцы будут определены как самцы. Такие определители обычно не ориентированы на переходные периоды (например, на период линьки), а при кольцевании работа может вестись как раз в это время.
МЕТОДЫ ПРИЖИЗНЕННОГО АНАЛИЗА ПТИЦ
Через руки кольцевателя проходит очень большое количество птиц, представляющих собой потенциально источник обильной информации, если ее правильно собирать. Важно, что эту информацию мы получаем сразу, и чем менее изучены территория или вид, тем она ценнее. При таком подходе изучение биологии птиц путем массового отлова, кольцевания и прижизненной обработки становится очень продуктивным методом для любой территории и любого вида.
У живой птицы можно измерять или оценивать многие, самые разнообразные показатели. Но результативность обработки птиц сильно возрастает, если изучают не набор случайных показателей, а определенную заранее продуманную систему их, построенную таким образом, чтобы получаемая информация была наибольшей и целенаправленной.
Системы анализа живых птиц разрабатывались в ряде руководств по кольцеванию (Baldwin et al., 1931; Witherby et al., 1938—1941; Cornwallis, Smith, 1960; Williamson, 1960—1964; Busse, Gromadzki, 1962; Svensson, 1970; Flegg, Zink, 1973).
Впервые близкую к современным систему анализа дополнительных признаков птиц предложил Маккаб (McCabe, 1943). Непосредственная цель этой системы была ориентирована на расширение сведений, приводимых на коллекционных этикетках. Но она оказалась удобной и для расширения данных, записываемых о птице во время кольцевания. Около пятнадцати лет назад эти данные стали рассматриваться как имеющие самостоятельную ценность, а кольцевание часто стало лишь сопутствовать сбору информации о живых птицах. За прошедшие годы усилиями многих исследователей методы прижизненной обработки птиц были отработаны, проверены и унифицированы. В наше время каждому исследователю нет необходимости придумывать свою систему, более того, это обычно и нежелательно, так как нарушает возможности сравнения и коллективных исследований.
В СССР впервые полная система обработки воробьиных птиц была применена на Биологической станции ЗИН АН СССР. Она была детально разработана и увязана с особенностями обработки птиц в других странах. Ниже приводится в качестве основной эта система; одновременно приведены основные варианты, встречающиеся у других исследователей.
Осмотр птицы сопровождают раздуванием перьев, в результате чего становятся видны линяющие перья, клоакальный выступ, наседное пятно и жировое депо под аптериями (см. рис. 3). Дуют равномерно, не сильно, под перо, под острым углом, от хвоста к голове. Одновременно поворачивают птицу, подставляя разные участки тела.
Скорость обработки птиц увеличивается, если операции разделены между двумя или тремя людьми, один из которых ведет запись в журнал.
Регистрируют следующие данные: номер кольца, название вида, пол, возраст, место поимки, год, месяц, число и час, длину крыла, вес тела, балл жировых резервов, стадию наседного пятна, форму клоакального выступа и размеры его у самцов, линьку и дополнительные данные, если они нужны. Для ускорения записи, ее компактности и облегчения чтения при обработке избегают словесной записи и пользуются цифровыми обозначениями и символами, вносимыми в соответствующие графы стандартного журнала. Вся операция в отношении нелиняющей птицы занимает 20—30 сек. При описании линьки — 1 —3 мин. После тренировки можно обрабатывать за день до 250 птиц. Для овладения приемами прижизненной обработки нужно несколько дней. Эта работа доступна студентам при соответствующем контроле.
Регистрируемые данные
Дата, час и место поимки. Эти данные обязательны и при кольцевании. Количество пойманных птиц. Абсолютных и строго объективных методов учета численности птиц или их миграции не существует. Поэтому всякий стандартно применяемый метод пригоден для определенных целей, имеет свои достоинства и недостатки. Подневное или суточное изменение количества пойманных стандартным методом птиц может характеризовать сроки и интенсивность перемещения их, дополняя данные, собранные другими методами.
Подвидовая принадлежность. В большинстве случаев различия между подвидами частично перекрываются индивидуальной изменчивостью. При определении по коллекционному материалу, к какому подвиду относится особь, используют и географический критерий, т. е. учитывают место добычи. Такой подход недопустим при кольцевании, так как здесь требуется точное и несомненное отнесение к тому или иному подвиду каждой особи. Выяснение подвидовой структуры вида — одна из задач кольцевания, которая решается анализом возвратов окольцованных птиц. Поэтому следует воздерживаться от записывания в журнал кольцевания тринарного названия птиц, исходя из географического положения места поимки. Но для облегчения будущего анализа желательно записывать, к какой из наблюдаемых вариаций относится птица, не придавая вариациям таксономического статуса. Позднее некоторые из этих вариаций могут оказаться принадлежащими птицам с особой гнездовой территорией, направлением или дальностью миграции.
Определение пола. У многих видов признаки пола известны, и определение не составляет проблемы. У других видов и в некоторых возрастах или нарядах оно затруднено. В первой половине лета форма клоакального выступа и наличие наседного пятна могут служить критериями пола у многих видов. Если определение пола сомнительно, его не следует вносить в запись. Вместо этого лучше в примечании записывать характер самого признака.
Впоследствии, когда станет ясно, надежен ли признак, эти пометки позволяют восстановить пол окольцованных птиц. Запись пола по непроверенным критериям, как бы они ни казались убедительными, навсегда вносит в первичную информацию неисправимые ошибки.
Определение возраста. К нему относятся все правила об определении пола. Кольцевание производится только после того, как сделаны перечисленные выше записи. Взвешивание. У летающих животных контроль веса тела осуществляется очень строго и все вариации веса неслучайны. Поэтому вес тела у птиц — более информативный показатель, чем у других групп животных.
Измерение длины крыла. Длина крыла — наиболее обоснованный линейный показатель размеров птицы в пределах одной группы — вида, популяции, пола. Так как фактически этот показатель отражает длину очень важного органа — пропеллирующей части крыла, случайные вариации его минимальны. Одно время сосуществовало несколько разных способов снятия промера, но теперь большинство международных руководств рекомендует применять только способ максимального выпрямления крыла (см. рис. 4). Преимущество этого способа — однозначность результатов. Кроме того, этот способ самый удобный при работе с живыми птицами.
Иные промеры. Для определения пола у некоторых видов измеряют длину хвоста; по длине хвоста слетков можно определять их возраст. Но в целом следует предостеречь от большого количества измерений. Как правило, такие измерения замедляют обработку, портят оперение кольцуемых птиц и, главное, никогда потом не обрабатываются, а обработанные обычно не дают биологически ценной информации.
Форма и размер клоакального выступа. Весной в период размножения это признак, позволяющий определять пол птицы, степень половой активности и стадию гнездового цикла (см. рис. 8).
На возможность использования клоакального выступа как критерия пола и степени половой активности впервые было указано Масоном (Mason, 1938) и Дростом (Drost, 1938), а позднее Волфсоном (Wolfson, 1954). В. Р. Дольник (1967) предложил метод полуколичественного определения половой активности самцов на основании двух промеров выступа и корреляции между произведением этих промеров и весом семенников. Степень увеличения клоакального выступа различна у разных видов, поэтому этот индекс непригоден для межвидовых сравнений, а корреляция между размером выступа и весом семенников должна быть установлена путем вскрытий для каждого вида отдельно.
У самца производят два промера клоаки — высоты выступа от прилежащих кпереди тканей брюшка и ширины его в средней, самой толстой части. Промеры записывают как произведение двух чисел.
Наседное пятно. На время размножения у самок всех видов воробьиных (а у части видов и у самцов) развиваются наседные пятна. В своем развитии наседное пятно проходит ряд стадий, удобных для регистрации (см. выше). Запись стадии наседного пятна производится в соответствующей графе римской цифрой, обозначающей номер стадии.
Пневматизация черепа. Птенцы вылетают из гнезд с непневматизированным черепом, пневматизация которого продолжается 2—7 месяцев. Важно научиться видеть пневматизацию черепа на живой птице (см. выше). Пневматизация черепа, как видно на рис. 14, условно подразделяется на пять различимых стадий (А, Б, С, D, Е). Запись производится в соответствующей графе одной латинской буквой, обозначающей стадию, или набором из двух-трех соседних букв, если установить ее точно не удалось.
Оценка величины жировых резервов. Подкожные жировые резервы изменяются коррелированно с изменением содержания жира в полости тела и тканях, всегда составляя половину общих запасов жира (Блюменталь, Дольник, 1962). Подкожный жир, расположенный в жировых депо, просвечивает через кожу и у мелких птиц виден при раздувании перьев. На этом основаны методы прижизненной оценки жирности птиц. В получившей всеобщее распространение в послевоенные годы системе оценки жирности (Блюменталь, Дольник, 1962) приняты следующие границы баллов.
«Нет» — нет видимых отложений жира ни на одном из основных проверяемых участков (межключичная ямка, боковые жировые депо под крылом, брюшко и жировые депо на крестце).
«Мало» — жир занимает менее половины поверхности депо.
«Средне» — жир занимает более половины депо, но не все.
«Много» — жир занимает все депо.
Тренированный человек может, вводя дополнительные баллы для промежуточных состояний, увеличить их число до 7—11, но, как правило, в этом нет необходимости.
Следует употреблять балльные оценки однообразно, всегда помня критерии границ баллов и одинаково применяя их ко всем видам, независимо от того, достигают ли в данном виде птицы высшего балла, или их жирность всегда ниже его (в этом случае для данного вида высшего балла просто не встречается).
Баллы «нет», «мало», «средне», «много» неудобно записывать словами, а писать их номерами не рекомендуется, чтобы избежать часто встречающейся ошибки — вычисления среднего балла для группы птиц путем сложения номеров баллов каждой птицы. Дело в том, что цена баллов, выраженная в граммах жира, неодинакова у разных баллов и их нельзя суммировать. Удобно обозначать баллы значками — символами, быстрыми в написании и легко отличимыми. В СССР получили распространение значки, предложенные Т. И. Блюменталь:
— — «нет»,
~ — «мало»,
+ — «средне»
X — «много».
Промежуточные баллы обозначаются добавлением знака < («меньше») и > («больше»); для предельно жирных птиц (балл «очень много») употребляется удвоенный знак: X X. Таким образом, между баллами «мало» и «средне», например, могут вставляться два промежуточных балла: > ~ и < +. Определяемое таким образом состояние птиц нужно называть жирностью, а не упитанностью, так как определяется именно количество жира. В методах определения упитанности оценивают одновременно и жирность и количество мышц, что не одно и то же.
Последний метод более применим в промыслово-гастрономической практике. Четырехбалльная система оценки жирности распространена в СССР, США и некоторых странах Европы. В Польше применяют семибалльную систему, которая, на наш взгляд, избыточна за счет более дробного деления птиц низкой жирности. Обе системы сравнивались между собой, и выработаны количественные приемы перевода данных,полученных одной системой, в другую (Busse, 1970).
Регистрация линьки. Линька — наиболее сложное для регистрации явление вследствие многоплановости процесса. В зависимости от задач она может быть очень детализирована или, наоборот, обобщена. Следует предостеречь от излишней детализации (если она не запланирована специально), так как опыт показывает, что слишком детальное описание линьки не поддается обобщениям и усреднениям при обработке результатов. В любом случае в первую очередь выделяют пять несомненных последовательных состояний смены одного пера: старое перо, выпавшее перо, трубочка, кисточка и новое перо. Эти состояния пера удобно записывать короткими символами. Широко применяется система символов, предложенная Т. И. Блюменталь. Достоинство этих символов — их мнемоническая связь с обликом перьев:
П — старое перо,
О — выпавшее перо,
| — трубочка (все перо в чехле),
\/ — кисточка (часть пера в чехле, а часть раскрылась),
/\ — новое, полностью освободившееся от чехла перо.
В отношении маховых и рулевых перьев стадию кисточки подразделяют на две:
V — перо не достигло 2/3 полной длины,
/\ — перо длиннее 2/3 полной длины.
Линьку полетных перьев (рулевых и маховых) записывают раздельно для каждого пера, указывая его номер. Всегда нужно проверять симметричность линьки на обоих крыльях и в обеих половинах хвоста.
При прижизненной обработке рекомендуется сквозная нумерация маховых от дисталыюй части крыла к телу.
Счет маховых удобно начинать с первого дистального длинного пера, называя его номером вторым.
Запись линьки маховых следует производить, перечисляя их номера сбоку от символов каждого состояния пера, например для первостепенных маховых: П1.2.3.4 O5 |6 V7 V8 /\9.10~.
Рулевые нумеруют центробежно; у воробьиных 6 пар рулевых. В разработанной Британским орнитологическим трестом и распространенной в Западной Европе методике записи линьки маховых и рулевых вместо символов состояния пера применяется нумерация состояний их:
0 — старое перо,
1 — отсутствует или трубочка,
2 — кисточка менее 1/3 полной длины пера,
3 — более 1/3 и менее 2/3,
4 — от 2/3 до полной длины,
5 — новое перо, без следов чехла.
Кроме того, цифра 8 означает перо полной длины, но новое или старое оно — не установлено, а цифра 9 — осмотр не производился. Такая система записи имеет свои преимущества, но она возможна, если для каждого пера в журнале есть специальная графа. В противном случае при чтении записи можно спутать номера перьев и номера состояний пера.
Линька оперения тела описывается раздельно по участкам. Участки могут соответствовать отдельным птерилиям, но это необходимо далеко не всегда. Обычно при массовых осмотрах достаточно трех участков: голова (так как она может начать линять позднее тела), спинная и брюшная стороны (последние разделены, потому что осматриваются отдельно).
Для каждого выбранного участка интенсивность линьки оценивается в баллах раздельно для каждой фазы смены пера (старые, трубочки, кисточки, новые). Применяется четырехбалльная оценка (нет, мало, средне, много). Границы баллов:
«нет» — совсем нет или лишь несколько перьев в данной фазе линьки (балл обозначается прочерком в соответствующей графе);
«мало» — кажется, что до 1/4 перьев в данной фазе (балл обозначается однократным написанием соответствующего фазе смены пера символа);
«средне» — кажется, что до 1/2 перьев в данной фазе (обозначение балла — двукратное написание символа);
«много» — более половины перьев в данной фазе (балл обозначается троекратным написанием символа).
Например, запись линьки для участка: ||VVV/\ означает, что старых перьев нет, трубочек средне, кисточек много и новых перьев мало.
Дополнительные признаки. У тех видов, цвет клюва которых имеет сезонные изменения, полезно записывать его, выделив последовательные стадии и присвоив им краткие символы. Это дает информацию о половом цикле. У видов, изменяющих в онтогенезе цвет глаз, полезно регистрировать его, что позволит найти критерии возраста.
При выкармливании птенцов у многих взрослых воробьиных, в том числе и у самцов (если они кормят), в углах рта вздуваются валики, отдаленно напоминающие птенцовые, но отличающиеся оранжевым цветом, сухостью и морщинистостью. Отмечая этот признак, можно выделять кормящих птиц. Форма клоаки самки позволяет безошибочно определить время спаривания и откладки яиц данной птицей. У самцов в период копуляции клоака также имеет особенности, позволяющие узнавать самцов в этом состоянии.
Формуляры записей
При накоплении массового материала о десятках тысяч осмотренных птиц, необходимости его обработки, в том числе и машинной, и важности обмена этим материалом выбор системы записи данных оказывается очень существенным. Почти одновременно и поэтому некоординированно развивались три системы записей; форма Британского орнитологического треста, форма польской Балтийской акции и распространившаяся в СССР форма, выработанная в Рыбачьем. Попытка стандартизировать формы записей, предпринятая Международным совещанием в 1971 г., не привела к выработке обязательной формы, хотя и был рекомендован один из компромиссных вариантов (см. Flegg, Zink, 1973).
К сожалению, этот вариант пока неудобен для большинства орнитологов СССР, так как он включает в себя ведение записи прямо в цифровом коде, рассчитанном на дальнейшую работу на электронных счетных машинах. В пределах СССР, видимо, пока можно придерживаться весьма распространенного стандарта, выработанного в Рыбачьем.
Применяются две формы ведения записей: журнальная и карточная. Карточная форма перспективнее в отношении дальнейшей обработки, но требует безупречно налаженного хранения и очень объемистая. Журнальная форма компактнее, удобнее при быстром ведении записей и надежнее сохраняется. Удобно использовать две формы журналов — для сезонов, когда нет линьки, и для сезона линьки. Во втором случае запись об одной птице помещается в одну строчку на страницу большого формата «лежа», 25 строк на странице. Во всех формулярах первые графы — обязательная при кольцевании информация: номер кольца, название вида, пол, возраст, дата, час, место, орудие лова, номер его, если их несколько. Далее идет информация, полученная при осмотре птицы. Сначала первостепенная: вес тела и длина крыла. Если делаются дополнительные промеры частей тела, относящиеся к третьестепенной информации, они могут быть поставлены за длиной крыла и объединены с ней общим надзаголовком «промеры». Далее следует информация, получаемая при раздувании перьев: балл жирности, стадия наседного пятна, размер клоакального выступа и линька. Последняя состоит из многих граф. Вслед за линькой идет третьестепенная информация — цвет клюва, глаз и т. п. Предпоследняя графа — пневматизация черепа, которую удобнее определять перед выпуском птицы, последняя графа — примечания.
Графы для записи линьки могут быть подразделены тремя способами, в зависимости от размеров журнала или карточки и задач исследования. Первый способ самый краткий и наименее удобный, под общим разделом «линька» три подраздела — «тело», «маховые» и «рулевые»; подраздел «тело» разделен на три графы — «голова», «спина» «брюхо». Размер каждой графы должен быть достаточен для написания в ней всего набора символов и номеров маховых и рулевых. Второй способ подобен первому, но подразделы «маховые» и «рулевые» содержат по пять граф, по графе для каждого состояния пера, от старого до нового. В эти графы записываются номера перьев в соответствующем состоянии.
Третий способ требует записи данных об одной птице на двух развернутых листах лежа. Подразделы «маховые» и «рулевые» разбиты каждый на число граф, соответствующих числу перьев в крыле (19 для воробьиных, 25 для неворобьиных) и в хвосте (в одной половине 6 для воробьиных и 8 для неворобьиных). Так как линька может быть асимметричной, если возможно, лучше делать эти графы «двухэтажными», помещая вверху запись о левом крыле, а внизу — о правом. В таком формуляре линька каждого пера заносится значком в графу, соответствующую номеру пера.
Этот способ записи самый надежный и быстрый; он рекомендован для международных исследований, но громоздкий. Наконец, для подробного изучения контурного пера и мелких перьев крыльев и хвоста и для них может вводиться разбивка на участки или птерилии. Такая полная запись размещается только на большой карточке, так как она занимает 80 граф. Чем подробнее запись, тем больше информации она содержит и тем проще поддается последующей обработке. С другой стороны, при осмотре птицы подробная запись линьки, особенно маховых и рулевых, происходит быстрее и, что особенно важно, приводит к меньшей вероятности ошибиться в нумерации маховых. Напротив, сокращенная запись требует большей тренировки и внимания при работе и сокращает объем полученной при осмотре птицы информации.
Опыт Биологической станции в Рыбачьем указывает на возможность компромиссного ведения записей линьки. Если исследователь осматривает мало линяющих птиц или намерен объединять свои данные с другими исследователями, например, для изучения географических вариаций в линьке, следует всех птиц описывать по подробной схеме. Если отлавливается очень много линяющих птиц, большую часть данных об их линьке можно записывать в компактной форме, но достаточную часть материала записать подробно. В отличие от записей в полевых дневниках журналы и карточки заполняют чернилами, так как это документы многократного пользования.
Регистрация данных повторных осмотров
Часть птиц попадается неоднократно, иногда по нескольку раз в день в течение целого сезона или даже несколько раз в течение ряда лет. Таких птиц нужно каждый раз подвергать повторному полному осмотру, так как они дают ценнейшую информацию об изменениях состояния конкретных птиц в естественных условиях.
Для повторных осмотров нужно иметь специальный журнал, а по окончании работ все данные о птице, включая первый осмотр, переносить на карточку.
Анализ погибших птиц
При отлове и кольцевании всегда какое-то количество птиц погибает. Их нужно обязательно использовать. В первую очередь производится описание птицы точно так же, как живой. Затем при вскрытии по пневматизации черепа и фабрициевой сумке точно определяют возраст, по гонадам — пол. Взвешивают гонады. Тело сохраняют в замороженном виде или фиксированным в спирте для последующей экстракции жира. В случае необходимости делают тушку. Эти птицы нужны для проверки применяемых критериев пола и возраста, поиска их, если они неизвестны, а также для определения реальной цены баллов визуальных оценок и для построения регрессий между внешними признаками и внутренними процессами.
КОРРЕЛЯЦИЯ ВНУТРИ СИСТЕМЫ АНАЛИЗИРУЕМЫХ ПРИЗНАКОВ
Перечисленных признаков, их изменений и записи достаточно для накопления полноценного исходного материала. Но для обработки этих данных необходимо ясно представлять себе связь - между признаками, сущность так называемых полуколичественных методов и способы нахождения конкретных параметров для этих методов.
Количество пойманных птиц
Данные о количестве пойманных птиц при применении стандартного метода отлова с успехом анализируются в нескольких направлениях и могут давать информацию, малодоступную для других методов.
Фенологические данные по срокам прилета и особенно отлета собираются этим методом в очень стандартной форме и часто лучше, чем при наблюдениях (см., например, Паевский, 1969, 1972).
Обнаружение орнитологических редкостей залетов сильно облегчается при отлове. Регулярные отловы хорошо фиксируют инвазии, их сроки и массовость. Анализ отловов незаменим при изучении сроков пролета половых и возрастных групп.
Изменения отловов очень точно фиксируют сроки начала летних перемещений, в том числе и отлета, почти не обнаруживаемые при визуальных наблюдениях (см., например, Блюменталь, 1971). Почасовые изменения отловов дают хорошие количественные представления о суточном ритме подвижности птиц разных видов и в разные сезоны.
Наконец, изменение числа пойманных птиц во время миграции отражает в отношении некоторых видов динамику перелета, сроки его начала и окончания. Для стандартизации этих данных, полученных в разных точках, теперь принята единая система суммирования их по пятидневкам (Flegg, Zirik, 1973). Чтобы пятидневки у разных авторов и в разные годы приходились на те же числа, предложена стандартизация их относительно календаря: год разбит на 73 пятидневки, начиная с 1 января. Двенадцатая пятидневка (с 25 февраля по 1 марта включительно) содержит в високосные годы 6 дней. Так как одни и те же подневные данные дают разную картину после объединения их по пятидневкам, начинающимся с разной даты, следует всегда придерживаться указанного стандарта независимо от даты начала отлова.
Вес тела и жирность
Вес тела сам по себе — признак с довольно расплывчатым значением, так как вес тела может зависеть от вариации истинных размеров птицы, вариации содержания в ней жира и воды. Чтобы стандартизировать данные о весе тела, в первую очередь нужно избавиться от влияния вариации жирности, то есть сравнивать веса птиц одинаковой жирности. При этом очевидно, что наиболее подходит для этих целей балл «нет», поскольку в пределах этого балла вариация жирности минимальна. Вес птиц с баллом жирности «нет» называется «тощим весом птиц» и рассматривается как первая, исходная для всех других построений, характеристика размеров особи, группы особей или вида. Поскольку скорость метаболизма в жировой ткани низкая, многие энергетические расчеты (расхода энергии в покое, энергии существования, потребления Нищи) производятся исходя из тощего веса. Только расход энергии на активность, в частности полет, производится по полетному (полному) весу тела.
Следующий этап — это освобождение данных о тощем весе тела от влияния вариации размеров. У большинства видов это достигается выравниванием данных по линейному размерному показателю — длине крыла. Обычно выравнивание приходится делать раздельно для полов, иногда возрастов и популяций. Длина крыла (линейный размер пропеллирующей части крыла) по аэродинамическим и энергетическим требованиям жестко связана с весом летательного аппарата. Корреляция носит степенной характер (Блюменталь, Дольник, 1970), но при коротком диапазоне вариации веса обычно с достаточной точностью аппроксимируется как линейная зависимость. По данным о длине крыла и тощем весе одних и тех же птиц вычисляется регрессия, по которой, зная размер крыла, можно вычислять соответствующий среднестатистический тощий вес и сравнивать его с реальным.
Тощий вес тела обладает сезонной вариацией, которая определяется вариацией содержания воды в теле и протеинов, оба показателя варьируют связанно и однозначно (Dolnik, 1973). Следовательно, обработка материалов должна производиться раздельно по сезонам, точнее по сезонным физиологическим состояниям, и одно из них должно быть выбрано опорным, стандартным. Наиболее подходит для этих целей период после завершения послебрачной или постювенильной линьки, но в случае необходимости может быть принят зимний период или первая половина периода весенней миграции (Блюменталь, Дольник, 1962). В эти сезоны тощий вес минимальный и наиболее стабильный по сравнению с периодами размножения и линьки. После того, как данные о весе птиц выровнены по размеру крыла путем прибавления (с соответствующим знаком) к реальному тощему весу коэффициента наклона регрессии на единицу длины крыла, умноженному на разность между средней для группы и реальной для данной птицы длинами крыла, полученные изменения тощего веса в течение времени отражают изменения содержания воды и протеинов в теле птицы. Так как колебания веса протеинов значительно меньше, чем колебания воды, то говорят только о колебании веса воды. Это очень важный показатель. Как выяснено, он отражает интенсивность процессов синтеза и утилизации протеинов и четко указывает на такие процессы, как рост, синтез яиц, подготовку к линьке или увеличение массы мышц перед миграцией или при зимовке в низких температурах. Во всех этих случаях тощий вес увеличивается. В непродуктивные сезоны года (зимой, осенью и ранней весной), вычитая из полного веса птиц рассчитанный для них по длине крыла тощий вес, получаем вес жировых депо (т. е. жира и сопутствующих накоплению жира веществ). Помимо перечисленных факторов на вес птиц влияет содержание пищи в пищеварительном тракте. Поэтому наиболее пригоден для всех сравнений вес пойманных ранним утром птиц. По этой же причине изменение веса птиц в течение дня может быть использовано для изучения суточных ритмов питания в природе и даже для косвенных расчетов расхода энергии в естественной обстановке. Если вычерчивается кривая изменения веса за сутки для всех пойманных птиц, то она не всегда будет отражать ритм питания по двум причинам: во-первых, в силу того, что любые методы отлова избирательны по отношению к птицам с разным поведением, а во-вторых, вследствие разных суточных ритмов поведения птиц в разном физиологическом состоянии. Например, в пределах светлой части суток максимум перемещений (но не локомоторной активности) птиц приходится на утренние часы, и в это время почти все способы отлова наиболее эффективны. Во время миграции в эти часы попадаются преимущественно более подвижные жирные птицы. Жирные и тощие птицы имеют разные ритмы питания и разный вес тела, поэтому их суммирование смазывает картину. Правильный метод — построение отдельных суточных графиков для тощих птиц и птиц с определенным баллом жирности. При таком построении обнаруживаются действительные ритмы питания. Для верных заключений о ритме питания исследователь обязательно должен учитывать особенности места отлова (трасса ли это интенсивного пролета или место остановки на кормежку, свойственная или не свойственная виду стация, водопой, место укрытия от дневного зноя и т. п.) и способа отлова (ловушка на летящих птиц, паутинная сеть, отлов на прикорм, водопой, ног манных птиц). Наиболее ценны исследования, в которых применялись разные методы отлова и разные стации.
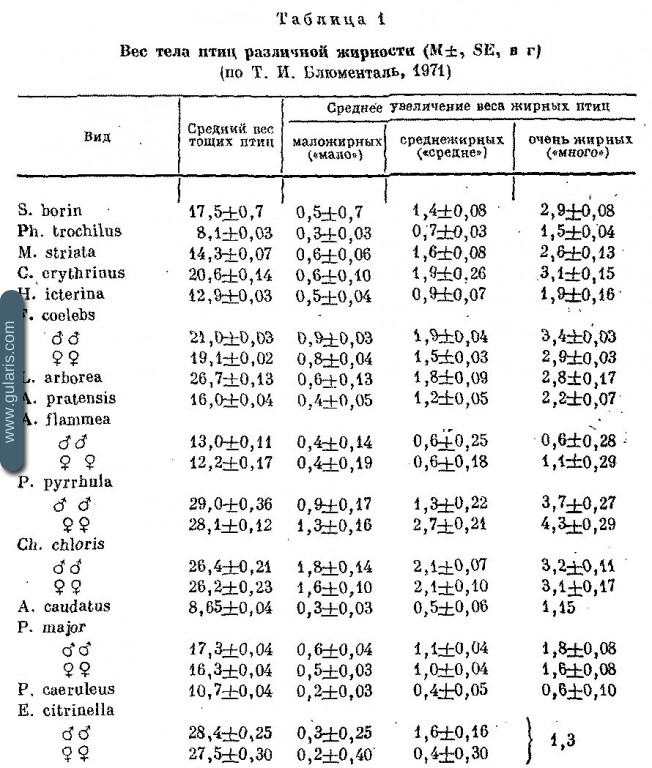
Средние веса птиц каждого балла жирности вычисляются по данным о весе птиц разных баллов жирности (табл. 1). Известные данные приведены и при повидовых описаниях признаков пола и возраста. Чтобы избавиться от влияния вариаций тощего веса, вычисление производится для самцов и самок раздельно и в пределах одного физиологического сезонного состояния. Различия в весе птиц разной жирности не полностью отражают изменения содержания жира в теле, так как параллельно отложению жира изменяется и вес нежирной части тела. Для точного определения цены баллов жирности в граммах жира используют погибших и фиксированных птиц.
Техника экстракции жира
Техника экстракции жира, применяемая в настоящее время орнитологами, стандартизирована, и ее не следует нарушать, так как результат зависит от применяемого для экстракции растворителя и методики. Ощипанную свежую или фиксированную в спирте тушку высушивают в термостате либо при 98° в воздушной среде, либо- (лучше) при 40° в вакуумном сушильном шкафу до установления постоянного веса, который записывают. После этого высушенную тушку размельчают в порошок в гомогенизаторе. Последний можно заменить кофейной мельницей или просто растереть тушку вручную в фарфоровой ступке. Гомогенизат переносят в пакетик из фильтровальной бумаги. Оставшийся на стенках жир собирают кусочками ваты сначала всухую, затем с эфиром, и помещают вместе с ватой в пакет, который закрывают, надписывают и высушивают до постоянного веса. Вес пакета записывают, а пакет помещают (сразу столько пакетов, сколько поместится) в аппарат Сокслета, где пробы в течение не менее 14 час. промываются циркулирующим петролейным эфиром. Окончание отгонки устанавливают пробой стекающего из сосуда с пакетами эфира. Капля эфира на чистом стекле после высушивания не должна оставлять радужного следа. По окончании экстракции пакеты извлекают, вновь высушивают до постоянного веса и сразу взвешивают. Разность в весе пакета до и после экстракции — вес жира в тушке. На каждый балл жирности нужно иметь 10—20 образцов, причем для высших баллов больше, чем для низших. Калорический эквивалент изменений веса тела устанавливают по изменению среднего веса птиц каждого балла и изменению их жирности (Дольник, Гаврилов, 1971). Калорический эквивалент позволяет производить расчет энергетического баланса по данным об изменениях среднего веса пойманных птиц и повторных взвешиваний одной и той же птицы. Для вычисления калорического эквивалента разность в цепе двух баллов жирности в граммах жира умножают на его калорийность (9,3 ккал/г) и делят на разность в цене тех же баллов по средним весам тела в граммах. Полученная величина (в ккал/г веса тела) используется при расчетах. Например, если в предмиграционный период средний утренний вес отлавливаемых птиц увеличивается на 0,3 г за сутки, а калорический эквивалент составляет 6,0 ккал/г, запасание энергии для миграции происходит со скоростью 1,8 ккал/сутки. Если птицы, разбившиеся о маяк на одном берегу моря, весят в среднем 25,0 г, а разбившиеся в те же дни на противоположном берегу 20,0 г, и известно, что это те же стаи, мигрирующие через море, то можно рассчитать расход энергии на перелет этой преграды и т. д.
Стадии размножения
Связь между стадиями наседного пятна и стадиями размножения выясняется путем отлова птиц на гнездах. При этом регистрируют, какой стадии наседного пятна соответствует каждая стадия гнездового цикла, и вариацию этих показателей. Когда связь установлена, открывается возможность определять у пойманной вне гнезда птицы ее положение относительно гнездового цикла, что при массовом отлове расширяет возможности изучения биологии вида.
Связь между размером клоакального выступа и весом семенников у самцов устанавливается по погибшим вскрытым особям. Строится корреляция между этими двумя показателями (отдельно для восходящей части кривых и для нисходящей). Когда корреляция построена, удается следить за изменением половой активности путем измерения у отлавливаемых птиц внешнего показателя. Для уточнения полового цикла иногда полезно использовать и такие критерии, как изменение цвета клюва и валиков в углах рта. По ходу потемнения клюва выделяются стадии, для каждой из которых выясняют корреляцию с весом семенников. Научившись узнавать размножающихся птиц, получаем возможность опознавать неразмножающихся особей в гнездовой период. Их процент в популяции и особенности поведения почти не изучены. Соотношения весов оперения на исследуемых участках тела (например, на дорзальной стороне, вентральной стороне, голове, маховых и рулевых) устанавливаются на погибших птицах. Перья выщипывают, собирают и взвешивают. Эти данные необходимы я нахождения коэффициентов к методике анализа линьки.
ПОЛУКОЛИЧЕСТВЕННЫЕ МЕТОДЫ
Накопление данных о баллах жирности птиц и описаний линьки само по себе не решает проблемы объективного анализа массового материала. Для этого необходимо перевести данные качественных, глазомерных оценок в такую форму, которая позволяет оперировать с этими данными обычными статистическими методами. Задача была решена разработкой так называемых «полуколичественных методов». Полуколичественный метод основан на глазомерном отнесении степени проявления признака у каждой особи к тому или иному баллу; затем при помощи дополнительных исследований объективными методами находят количественное выражение среднего значения признака и границы вариации; баллам присваиваются номера, соотношение которых пропорционально среднему значению балла, найденному объективным методом. Далее с этими номерами баллов поступают, как с обычными количественными признаками,— вычисляют по ним средние значения для выборок, статистические ошибки, уровень достоверности различия и т. д.
Полуколичественный метод — это статистический метод. Он действует как количественный, если выборка достаточно велика. В отношении каждого отдельного определения это качественный метод. Если мы возьмем одного зяблика и на основании осмотра отнесем его жирность к баллу «средне» (цена балла 1,6 г жира), мы не можем утверждать, что этот зяблик содержит 1,6 г жира. Реальное содержание жира в нем (если мы верно отнесли его к данному баллу) может быть от 0,9 до 2,5 г. Таков диапазон вариации результатов экстрагирования жира из зябликов, отнесенных к баллу «средне». Но если мы имеем группу из 100 зябликов, отнесенных к баллу «средне», то средняя жирность этих птиц составит 1,6 ± 0,05 г, т. е. в отношении группы мы можем пользоваться визуальной балльной оценкой как количественным признаком. Разработка коэффициентов полуколичественного метода должна производиться заново для каждого вида, а при более тонких исследованиях'и для каждой популяции. Она проходит три последовательных этапа. Первый этап — глазомерная оценка широты вариации признака и разбивка его на ограниченное число баллов. Даже если изменчивость признака непрерывна, глаз человека может найти несколько устойчивых условных границ между баллами. Ошибка глазомерной оценки у тренированного человека при четырех- или пятибалльной системе не выходит за пределы ± 0 , 5 балла. Если критерий балла выбран четко, то расхождения в оценке признака независимыми исследователями практически не бывает. На этом этапе баллам присваиваются словесные названия и графические символы и начинается сбор материала.
Второй этап — определение среднего количественного значения признака, которому соответствует данный балл. Путем измерения количественными методами (например, экстракцией жира, подсчетом и взвешиванием перьев и т. д.) устанавливается среднее значение балла, производится статистический анализ вариации количества признака в пределах балла и достоверности средних значений. На этом же этапе в зависимости от того, какой точности задачу собираются решить, рассчитывают необходимый объем материала, исходя из размаха вариации признака и предполагаемой разницы в средних значениях, которая должна быть достоверной с тем или иным доверительным уровнем. Третий этап — присвоение баллам номеров пропорционально среднему численному значению признака. Например, установлено, что у данного вида баллу «нет» соответствует содержание 0,4 г жира в теле; баллу «мало» 0,9; баллу «средне» 1,6 и баллу «много» 2,5 г жира. Тогда номера баллов удобно обозначить как 4, 9, 16, 25. Если числа получаются не целые, их целесообразно округлить в разумных пределах. В результате появляется возможность суммировать балльные оценки отдельных птиц, вычислять средние и производить все расчеты обычными статистическими способами. В тех случаях, когда признак оценивается просто и удобно, иногда при окончательном представлении результатов его можно выражать не в не имеющих наименований значениях баллов, а прямо в наименованиях самого критерия, указывая при этом, что речь идет о полуколичественном методе. Так, среднюю жирность группы птиц можно выразить баллом 12,7, а можно сказать, что она составляла 1,27 г в весовом выражении визуальной оценки жирности. Лучше всего выражать баллы жирности через вес жира. Но это требует определения средней цены баллов жирности путем экстракции жира. Если исследователь не располагает возможностью для экстракции жира, или исследует сразу много видов и экстракция жира из каждого из них требует неоправданно большого количества птиц, или сам характер исследования не связан с биоэнергетическими оценками, можно ограничиться определением соотношения баллов весовым методом. Предположим, что самки данного вида при балле «нет» весят в среднем 20,5 г; при балле «мало» 21,0 г; при балле «средне» 22,0 г; и при балле «много» 24,5 г. В таком случае первому баллу присваивается номер 0 (так как мы не знаем, сколько жира содержится в теле нежирных птиц этого вида). Второй балл будет иметь номер 21,0—20,5 = 0,5. Третий балл соответственно 1,5, четвертый балл —4,0. Ценой балла, таким образом, оказывается превышение в граммах среднего веса птиц каждого балла над весом птиц с баллом «нет». Для удобства подсчетов в данном случае баллы удобно перевести в целые числа (0, 5, 15, 40), а в некоторых случаях — округлить.
Клоакальный выступ изменяется по каждому промеру за период половой активности в 5—10 раз, в то время как вес семенников в 200—600 раз. Поэтому в качестве индекса изменения размеров клоакального выступа удобно брать не изменение одного промера, а произведение промеров. По методу Дольника (1967), в качестве индекса используется произведение высоты выступа на его ширину (этот показатель изменяется в течение половой активности в 100 раз). Но можно использовать и произведение трех промеров — высоты и двух промеров по ширине, т. е. условного объема выступа. Тогда показатель будет варьировать почти в 1000 раз. В большинстве случаев это неудобно, так как затрудняет графическое изображение процесса изменения размеров выступа в линейных координатах.
АНАЛИЗ ДАННЫХ О ЛИНЬКЕ
Линька наиболее сложна для количественного анализа по массовому материалу вследствие того, что линька включает в себя много отдельных интересных процессов, которые кажется невозможным суммировать без ущерба для полноты картины. Поэтому разработано несколько методов анализа данных о линьке, каждый из которых исследует одну из ее сторон.
Птерилография. Если неизвестно, полная или частичная линька у данного вида или популяции, исследование линьки начинается с птерилографии. Для этого описания линьки у части пойманных птиц производятся раздельно для всех или большинства птерилий, особенно для отдельных групп кроющих крыла, в линьке которых наблюдается большая изменчивость. В специальной статье Г. А. Носкова и А. Р. Гагинской (1972) предложен хороший метод выделения описываемых участков. Впоследствии, при обработке материала, сравнивая описания разных птиц в разное время и особенно повторные описания окольцованных птиц, реконструируют сроки и полноту линьки на каждом участке. Полученная схема линьки, во-первых, упрощает обработку данных в других направлениях, а во-вторых, если сроки смены перьев на разных участках отличаются, позволяет создать таблицу стадий линьки, что очень важно при изучении неполной постювенильной линьки.
Выделение стадий линьки маховых. Два метода выделения стадий линьки маховых оправдали себя и получили распространение — это метод Блюменталь и Дольника (1966), основанный на выделении 11 стадий по номерам маховых и порядку их смены и метод Британского орнитологического общества (Evans, 1966; Newton, 1966; Snow, 1967), основанный на определении числа сменившихся маховых. Достоинство второго метода в том, что его можно применять к любым видам, ничего предварительно не зная о их линьке. Недостаток — невозможность понять по номеру стадии, в каком состоянии линьки отдельных маховых была птица. Этот недостаток отсутствует в первом методе, так как ключом к определению номера стадии служит определенное состояние линьки каждого махового. Но зато предварительно необходимо проверить, соответствует ли у данного вида линька схеме, взятой за основу ключа. Целесообразно использовать оба метода, поэтому ниже они оба описаны.
П е р в ы й метод. Путем сравнения записи линьки маховых с ключом, приведенным в табл. 2, находят стадию линьки.

Линька конкретной птицы может быть описана на промежуточной стадии. При бурной линьке могут совмещаться две или даже три соседние стадии. Таким образом, определение стадии требует квалификации и некоторого умственного усилия. Если в описание линьки птицы вкралась ошибка в нумерации маховых, ее обычно удается обнаружить, так как первостепенные маховые у воробьиных сменяются центробежно, от границы между второстепенными и первостепенными, а второстепенные маховые—центростремительно, с двух концов ряда.
Приведенным ключом можно пользоваться для всех воробьиных, исключая Muscicapa striata, но лучше, если по данным о линьке в изучаемой популяции для нее составлен свой особый ключ. Этот ключ одновременно служит и наглядной схемой порядка линьки маховых у вида или популяции, т. е. несет важную биологическую информацию.
В т о р о й м е т о д . Не требует ключа и нумерации перьев. Из исходной записи устанавливается, сколько было в крыле старых перьев, выпавших, трубочек, кисточек менее 2/3 длины полюго пера, кисточек более 2/3 его длины и новых перьев. Этим пяти состояниям пера присвоены баллы: 0 — старое, 1 — выпавшее перо, 2 — трубочка, 3 — малая кисточка, 4 — большая кисточка, 5— новое перо. Для определения стадии линьки число перьев в каждом из пяти состояний умножают на балл этого состояния, произведения суммируют. Стадия 0 — нет линьки, стадия окончания линьки равна упятеренному числу перьев в крыле. Соответственно при 19 маховых она равна 95. Иногда стадии линьки определяют раздельно для первостепенных маховых, второстепенных маховых и рулевых.
Графическое изображение линьки маховых. Как в первом, так и во втором методе при нормальной скорости линьки время прохождения каждой стадии одинаково, поэтому изображение линька на графике зависимости стадий от времени образует прямую линию. Эта особенность и составляет ценность методов. Редко когда удается проследить всю линьку в популяции от начала до конца (хотя бы потому, что во второй половине ее птицы начинают откочевывать). Метод изображения линьки в форме линейного процесса позволяет по двум данным, полученным в разное время рассчитать срок начала и окончания линьки (если скорость ее не изменялась). Практически это делается двумя способами. В первом рассчитывают среднюю стадию линьки выборок птиц, пойманных в каждую из декад. По полученным точкам рассчитывают регрессию, из которой находят даты начала и окончания линьки в популяции. При втором способе на график наносят точки, соответствующие повторным осмотрам одних и тех же окольцованных птиц, и данные, касающиеся каждой птицы, соединяются линиями. Получается наглядная картина протекания линьки в популяции.
Индекс регенерации оперения. Главным обобщающим критерием скорости линьки и изменения этой скорости может служить индекс регенерации оперения, расчет которого разработан применительно к воробьиным (зяблику, в частности) В. Р. Дольником и В. М. Гавриловым (1974). В основе метода лежит нахождение средних численных соотношений числа линяющих перьев для каждого балла интенсивности линьки на обследуемом участке, соотношения общего веса оперения на каждом участке и скорости регенерационных процессов (синтеза вещества) на разных стадиях роста пера. Подсчет реального числа линяющих в одной фазе перьев на птицах, отнесенных к каждому из четырех баллов, показал, что в среднем баллу «мало» соответствует ситуация, когда в данной фазе линьки находится 20% перьев; баллу «средне» - 40% перьев, и баллу «много» — 60% перьев. Отсюда соотношении баллов принято соответственно: 0 для балла «нет», 1 — для балла «мало», 2 — для «средне» и 3 — для «много».
Взвешивание общего количества перьев на трех основных участках дало следующее соотношение (весовые эквиваленты) 1 (перья головы), 4 (оперение спины), 5. (полетные перья) и I (оперение брюшной стороны). Интенсивность синтеза веществ) в пере оказалась в фазе кисточки в 4 раза ниже, чем в фазе тру бочки, она принята за 1 для трубочки и за 0,25 для кисточки. Для вычисления индекса общей массы регенерирующих перье] числовое значение баллов кисточек и трубочек на каждом участке умножают на весовые эквиваленты масс перьев на этих участках. Для первостепенных и второстепенных маховых и рулевых весовой эквивалент одного пера принят равным 0,10, так как общее число этих перьев 48 или 50, а их суммарный весовой эквивалент — 5.
Результаты по всём отдельно обследовавшимся участкам суммируют раздельно для трубочек и кисточек. Индекс регенерации оперения есть сумма полученного выше индекса общей массы перьев в фазе трубочек плюс 0,25 индекса общей массы оперения в фазе кисточек. Индекс регенерации оперения отражает интенсивность синтеза кератина и формирования перьев в течение линьки.
Показано, что интенсивность линьки строго коррелирует с изменением содержания воды в теле. Поэтому в качестве показателя интенсивности линьки в популяции можно использовать и изменение тощего веса птиц. Стадии линьки при неполной линьке (когда не сменяются маховые) представляют собой нерешенную проблему. В сущности, их можно выделять только по различию в сроках начала и окончания линьки на разных птерилиях. Дольник и Гаврилов (1974) составили в качестве первого опыта ключевую таблицу для определения стадий линьки у молодых зябликов, однако пользование ею требует значительной тренировки и не все стадии удается определить. В сущности, линька перьев тела хорошо разделяется на три периода: «начало» — от начала смены оперения до начала линьки перьев головы; «середина» — на всем теле много трубочек и кисточек, линяет голова; «окончание» — на теле и голове мало трубочек и много кисточек и новых перьев.
* * *
При хорошо налаженном регулярном отлове птиц метод прижизненного анализа оказался самым эффективным методом изучения биологии вида или популяции. Сроки миграции разных половых и возрастных групп и состояние мигрирующих птиц, летние перемещения, занятие и покидание территории, суточные ритмы активность и связь этих процессов с внешними факторами прекрасно устанавливаются при применении стандартных методов отлова. Метод дает обширные данные о сроках гнездования птиц, индивидуальной вариации сроков, повторном размножении, участии полов в насиживании и выкармливании, изменениях половой активности и поведении неразмножающихся птиц. Линька может быть изучена всесторонне, подробно и на большом материале, в том числе и линька одних и тех же птиц. Все процессы предмиграционного периода, включая суточные ритмы питания и отложения жира, выступают очень наглядно.
В течение миграции прослеживается энергетика перелета, состав мигрирующих стай, структура волны пролета как результат взаимодействия различий в поведении жирных и тощих птиц с внешними условиями. В зимний период вскрывается зависимость веса птиц от температуры среды, длины кормового дня и других факторов.
Методом прижизненной обработки выявляются и многие явления физиологического порядка. Все это делает работу полевого орнитолога эффективной, полноценной и интересной, а кольцевание птиц — оправданным независимо от того, скоро ли и в каком количестве будут получены сообщения о находках окольцованных птиц.
АКАДЕМИЯ НАУК СССР ЗООЛОГИЧЕСКИЙ ИНСТИТУТ Н. В. Виноградова, В. Р. Дольник, В. Д. Ефремов, В. А. Паевский Справочник. М., «Наука», 1976.- Войдите на сайт для отправки комментариев
















По многочисленным заявкам, привожу основополагающее руководство с картинками: как и зачем мерить птиц, и многое другое что помогает изучать птиц на новом, детальном и более высоком уровне, чем это было возможно до этого.
Обратите внимание существует некоторое разхождение в топографических названиях групп перьев, с те что я привел в "Лирическом отступлении". Некоторые детали я не учел, но намеренное применение именно той топографии, что приведено у меня - значительно полней подходит для описания снегирей.